Prostate epithelial cells are plated into mouse organoid culture where they form organoids, which are harvested prior to preparation for downstream analysis (Figure 1).
Basal and luminal epithelial cells are isolated using FACS. After excluding DAPI+ cells and depleting Lin+ cells (CD45, CD31, Ter119), basal and luminal cells are distinguished based on differential expression of EpCAM and CD49f (Figure 2). The approach described to plate prostate basal and luminal cells into organoid culture entails: (1) plating cells into matrix gel rings, and (2) coating wells with Poly-HEMA. Plating into rings has been previously described in Agarwal et al9. Utilizing this approach (Figure 3A) allows researchers to more easily avoid the matrix gel while replenishing the media (Step 3), and more easily count organoids by following the circumference of the well. Coating wells with Poly-HEMA has been shown to prevent 2D colony formation in retinal organoids17; however, this approach has not been utilized in the prostate organoid model. Importantly, coating wells with Poly-HEMA (Table 3) eliminates the occurrence of 2D colonies without interfering with organoid formation (Figure 3B). These modifications expand the capabilities of the prostate organoid assay.
Basal and luminal cells form organoids with distinct morphologies (Figure 4A). While most basal-derived organoids are similar in size (100-300 µm diameter) after 7 days in culture, luminal-derived organoids exhibit significant heterogeneity (30-450 µm diameter). Furthermore, most basal-derived organoids contain lumens surrounded by multi-layered epithelium (Figure 4A, top), whereas luminal-derived organoids range in morphology from hollow, with single-layered epithelium to solid, with multi-layered cords of cells that do not canalize (Figure 4A, bottom). The approaches described above to prepare organoids for downstream analysis (Steps 4, 5), were used to investigate whether these phenotypic differences are reflective of differences in lineage marker expression. Western blot analysis revealed that basal and luminal-derived organoids retain features associated with basal and luminal primary cells. Basal-derived organoids express higher levels of the basal marker cytokeratin 5 (K5), whereas luminal-derived organoids express higher levels of the luminal marker cytokeratin 8 (K8) (Figure 4B). Both basal and luminal markers were detected in basal and luminal-derived organoids in the bulk population, perhaps suggestive of differentiation (Figure 4B).
We sought to characterize lineage marker expression in basal-derived organoids and determine whether morphologically distinct luminal-derived organoids exhibit differences in marker expression by staining intact organoids and performing confocal microscopy (Figure 4D). Basal-derived organoids contained multi-layered epithelium with outer layers expressing high levels of the basal marker p63 and moderate levels of the luminal marker K8 (p63hi, K8mid), and inner layers without detectable levels of p63 and high levels of K8 (p63lo, K8hi) (Figure 4D, top). While all cells in single-layered luminal-derived organoids stained positively for K8, only select cells contained nuclear p63 (Figure 4D, bottom). These data validate the approaches to harvest and prepare organoids for analysis by Western blot or confocal microscopy and thereby expand the capability of the organoid assay to study key cellular processes, including differentiation.
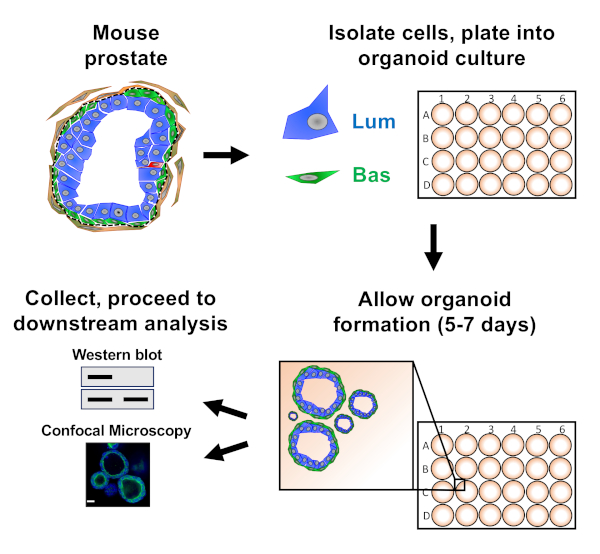
Figure 1: Schematic illustrating workflow to generate prostate organoids for collection and analysis. Total mouse prostate is dissociated and basal and luminal prostate epithelial cells are isolated by fluorescence-activated cell sorting via established protocols18,19. Basal or luminal cells suspended in a mixture of mouse organoid media and matrix gel are plated into matrix gel rings. After 5 to 7 days of culture, organoids are harvested for analysis by Western blot or confocal microscopy. Please click here to view a larger version of this figure.
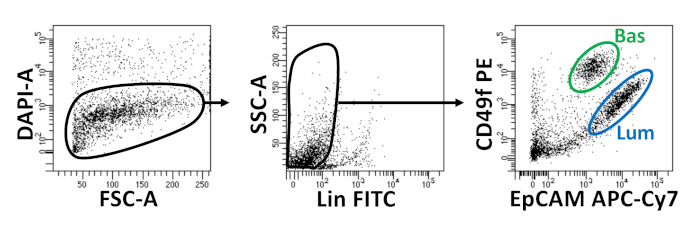
Figure 2: Isolation of mouse basal and luminal prostate epithelial cells using fluorescence-activated cell sorting (FACS). Dissociated cells from mouse prostate are stained with DAPI, to distinguish live from dead cells, and surface antibodies, to distinguish basal from luminal cells, prior to FACS. Left = Gated on DAPI- cells. FSC-A = forward-scatter. Center = Gated on Lin– cells (CD45lo, CD31lo, Ter119lo). SSC-A = side-scatter. Right = Basal cells (Bas) (EpCAMhi, CD49fhi), Luminal cells (Lum) (EpCAMhi, CD49fmid). Please click here to view a larger version of this figure.

Figure 3: Establishment of mouse prostate organoids. (A) Schematic illustrating approach to generate a matrix gel ring in a well of a 24-well plate. (B) Representative phase contrast images of organoids (3D growth plane) and two-dimensional colonies (2D growth plane) formed 7 days after plating prostate epithelial cells into un-coated (Poly-HEMA (-)), or coated (Poly-HEMA (+)) 24-well plates. Boxed regions within 2D growth plane are magnified on the right. Scale bars = 200 µm. Please click here to view a larger version of this figure.
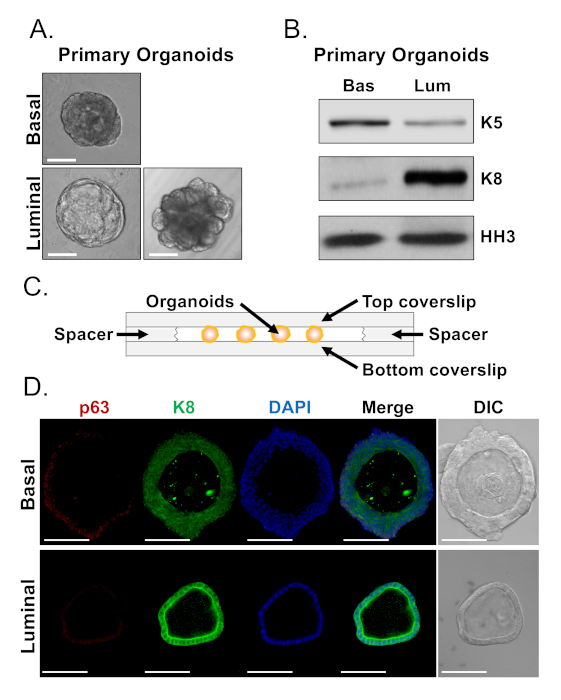
Figure 4: Analysis of lineage marker expression in prostate organoids by Western blot and whole-mount confocal microscopy. (A) Representative phase contrast images of basal-derived (top), and luminal-derived (bottom) organoids after 7 days of culture. Scale bar = 100 µm. (B) Western blot analysis of basal-derived (Bas) and luminal-derived (Lum) organoids after 5 days of culture. Staining for the basal marker, cytokeratin 5 (K5), and the luminal marker, cytokeratin 8 (K8), and a loading control, histone H3 (HH3). (C) Schematic illustrating chambered coverslip with spacers. (D) Representative differential interference contrast (DIC) and immunofluorescent images of basal-derived (top) and luminal-derived (bottom) organoids after 7 days of culture. Staining for p63 (red), K8 (green) and DAPI (blue) individually and merged. Scale bars = 100 µm. Please click here to view a larger version of this figure.
| Recipes | |
| Dispase-containing media | 1 mg/mL dispase + 10 µM ROCK inhibitor in advanced DMEM F12. Filter sterilize using 0.22 µm filter. |
| Dissociation media | 10% FBS + 1x Penicillin-Streptomycin in RPMI 1640. Filter sterilize using 0.22 µm filter. |
| Protein lysis buffer | RIPA buffer + phosphatase inhibitors + protease inhibitors |
| Blocking solution | 10% FBS in PBS with 0.2% Triton X-100 |
Table 1: Instructions for the preparation of key solutions.
| Component | Concentration |
| B-27 | 1x (dilute from 50x concentrate) |
| GlutaMAX | 1x (dilute from 100x concentrate) |
| N-acetyl-L-cysteine | 1.25 mM |
| Normocin | 50 µg/mL |
| Recombinant Human EGF, Animal-Free | 50 ng/mL |
| Recombinant Human Noggin | 100 ng/mL |
| R-spondin 1-conditioned media | 10% conditioned media |
| A83-01 | 200 nM |
| DHT | 1 nM |
| Y-27632 dihydrochloride (ROCK inhibitor) | 10 µM |
| Advanced DMEM/F-12 | Base media |
| R-spondin 1-conditioned media is generated as described in Drost, et al.13. After addition of all components, filter sterilize mouse organoid media using 0.22 µm filter. ROCK inhibitor is only added during establishment of culture and passaging of organoids. | |
Table 2: Instructions for the preparation of mouse organoid media.
| Protocol for preparing Poly-HEMA-coated plates | |
| 1 | Add 0.25 g Poly-HEMA to 50 mL 98% EtOH. Dissolve Poly-HEMA at 37 °C on a shaker. This process takes at least 4 h. |
| 2 | Filter sterilize Poly-HEMA using 0.22 µm filter. |
| 3 | Add 200 µl of Poly-HEMA solution per well of a 24-well plate(s). |
| 4 | Remove lid(s) from 24-well plate(s) after adding Poly-HEMA and allow solution to evaporate overnight. |
| 5 | Wash each well twice with PBS and ensure wells are completely dry prior to storage after final wash. NOTE: Disrupting the Poly-HEMA coating during washing could contribute to 2-dimensional growth upon plating epithelial cells into organoid culture. To prevent damage to Poly-HEMA-coated wells, avoid direct contact with the pipette tip while washing. The integrity of the Poly-HEMA-coated wells will remain intact unless the Poly-HEMA is scraped off by the pipette tip. |
| 6 | Poly-HEMA-coated plates can be stored at 4 °C for up to two weeks. NOTE: Wrapping plates in parafilm prior to storage will reduce the risk of contamination. |
Table 3: Protocol for preparation of Poly-HEMA-coated plates.
