I 1901 innførte den nederlandske botaniker Hugo de Vries betegnelsen mutasjon1. Twenty-seks år senere, da Hermann Joseph Muller oppdaget mutagent handling av røntgenstråler2, var mutasjoner allerede oppfattet som en av drivkreftene i evolusjon. Imidlertid var innholdet av mutasjoner ikke klart. For å besvare det fundamentale spørsmålet om mutasjoner oppstår spontant (dvs. en spontan mutasjon) eller som svar på valg (dvs. en indusert mutasjon), en metode var nødvendig for å observere mutational hendelser. En slik metode vil måle forventet antall mutasjoner per celle divisjon eller det som allerede var kjent som en mutasjon rate3,4.
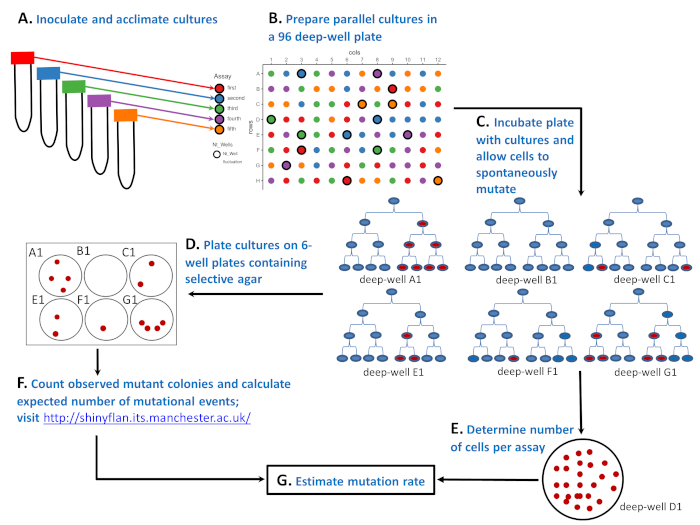
Figur 1: skjematisk illustrasjon av hvordan du utfører svingningene analysen med en mikrobiell belastning i en 96 dyp brønn plate. (A) vaksinere og acclimate celler i 50 ml rør som inneholder fem forskjellige miljøer (‘ rød ‘, ‘ blå ‘, ‘ grønn ‘, ‘ lilla ‘ og ‘ oransje ‘ analyser). (B) forberede parallelle kulturer med et lite antall følsomme celler i en 96 dyp brønn plate. Den “røde” analysen har 20 parallelle kulturer, mens den “blå”, “grønn”, “lilla”, og “oransje” analysene alle har 19 parallelle kulturer. Plasseringen av parallelle kulturer på 96 dyp brønn plate er tilfeldig. Tilfeldiggjøring kan gjøres med utfyllende LayoutGenerator. R skript eller av et annet verktøy. Oppsettet øverst til høyre er et tilfeldig resultat. (C) ruge den 96 dyp brønn plate og la cellene til å dele og spontant mutere. Seks kulturer fra dype brønner a1, B1, C1, E1, F1 og G1 viser hvordan antall mutanter svinger: 4, 0, 2, 2, 1, og 4 røde celler etter tredje celle divisjon, henholdsvis. Antall mutanter skiller seg ikke bare på grunn av ulike antall spontane mutasjoner (0, 1, eller 2 som vist ved den første røde cellen), men også fordi det er viktig når under en kultur syklus en motstand mutasjon spontant framgår (celle divisjon 1, 2, eller 3). (D) etter inkubasjons av 96 dyp brønn plate antall mutanter bestemmes av plating 81 parallelle kulturer. På oppsettet er disse sirkler uten uthevet kanter. Hele parallelle kulturen er belagt på en brønn av en 6 brønn plate som inneholder en selektiv agar. (E) de resterende 15 kulturene er fortynnet og belagt på ikke-selektiv agar å bestemme gjennomsnittlig antall celler (Nt). På layout disse er merket som Ntwells og har uthevet kanter. For hver analysen Nt er i gjennomsnitt over tre parallelle kulturer. Nederst til høyre er en Petri parabolen inneholder en ikke-selektiv agar plate med 25 CFUs av en utvannet kultur dyrket i en dyp brønn D1 (del av en “grønn” analysen). (F) etter inkubasjons av de selektive 6 brønn platene ble antallet observerte mutanter telt, og det forventede antallet mutational hendelser, m, ble estimert ved hjelp av et estimat for maksimal sannsynlighet. (G) å vite både antall mutasjoner, m, og antall celler per analyse, Nt, mutasjon raten ble anslått som m/Nt. Vennligst klikk her for å se en større versjon av dette tallet.
Salvador Luria og Max Delbrück i 1943 ga en genial løsning på dette problemet med svingninger analysen5 (se figur 1). Analysen starter med flere populasjoner (navngitte parallelle kulturer) som er initiert med et lite antall mikrobielle celler (figur 1a, B). Etter vekst i et godartet, ikke-selektiv miljø (figur 1C), parallelle kulturer er overført på plater som inneholder en selektiv markør (phages, antibiotika, etc.), der bare celler med en motstand mutasjon overlever og kan produsere en koloni (figur 1D). Den største forventningen var at hvis motstanden mutasjoner er indusert, antall celler som bærer en mutasjon bør fordeles mellom ulike populasjoner med gjennomsnittet lik variansen. Hva Luria og Delbrück funnet med svingninger analysen er at antall mutanter svingt drastisk og at variansen i antall mutanter blant ulike populasjoner var betydelig større enn gjennomsnittet. Luria og Delbrück viste dermed at mutasjoner er spontane. De viste at mutasjoner spontant dukker opp når DNA er kopiert, og antall mutanter avhenger av når mutasjonen oppstår under veksten av befolkningen. Se figur 1C, hvor seks populasjoner, hver startet med en mikrobiell celle (i blått), opplever ingen, 1 eller 2 enkelt mutasjoner. Populasjoner a1, E1, og F1 opplevde en enkelt mutasjon (første røde celle), men fordi en enkelt mutasjon spontant fremkommer på ulike tidspunkt under en kultur syklus, befolkninger endte opp med et helt annet antall observerte mutanter (fire, to og en, henholdsvis). På den annen side, bestander C1 og G1 endte opp med det samme antall observerte mutanter som E1 og a1, til tross opplever to mutational hendelser snarere enn én. Svingningene i observert mutanter blant populasjoner ikke bare ga analysen navnet, men viste også at en mutant frekvens (dvs. andelen av muterte celler) er en utilstrekkelig indikator på mutasjon rate.
Det overordnede målet for svingningene analysen er å estimere den spontane mutasjon rate av en bestemt genotype av bakterier eller andre encellede organisme vokser i et bestemt flytende miljø. Svingningene analysen er fortsatt den mest hensiktsmessige verktøy for å studere miljømessige avhengigheten av mikrobiell mutasjon priser og gir rask og rimelig mutasjon rate estimering. Alternative tilnærminger til mutasjon rate estimering, for eksempel maksimal dybde sekvensering6, befolkning sekvensering7, mutasjon akkumulering eksperimenter8, eller sammenligne Genova sekvenser av et avkom til de av foreldrene9 er mye mer arbeidskrevende, og dermed dårlig egnet til potensielt oppdage miljømessige avhengigheter. Men dynamiske aspekter ved en mutasjon generasjon og reparasjon er i stor grad utilgjengelige for en svingninger analysen eller til noen av metodene for assaying en mutasjon rate nevnt ovenfor. For å studere hvordan antall mutasjoner endres i tid, rom, eller blant individuelle celler i en populasjon, tilnærminger én celle11,12 er nødvendig, som i tillegg til å være mer arbeidskrevende enn svingninger analyser, krever høyt spesialiserte ferdigheter og utstyr.
I praksis er en svingninger analysen teller celler få en fenotypiske markør på grunn av en mutasjon som oppstår i et miljø som mangler valg for at markøren. Den meta-analyse av hundrevis av publiserte analyser10 viser at minst 39 forskjellige fenotypiske markører har blitt brukt siden analysen er starten i 1943. Svingningene analysen kan brukes til å sammenligne gjennomsnitt og miljømessige avhengighet av mutasjon priser blant laboratorium, klinisk, nonmutator, og Mutator stammer vokser i ettergivende miljøer. Analysen gjør det mulig for mutasjon rate estimering i celler med ulik genetisk bakgrunn vokser i enten minimal eller rike miljøer. Analysen er egnet ikke bare for populasjoner som vokser som en monokultur, men kan også brukes til å studere virkningene av celle-celle interaksjoner på mutasjon priser11. Når belastningen av interesse er cocultured med en annen belastning, og en nøytral markør brukes til å skille stammene, mutasjon priser kan analyseres for to stammer i samme rør samtidig.
Svingninger analyser har avdekket at den spontane mutasjon rate avhenger av både en celle genotype og dets miljø12 og er en egenskap som selv utvikler seg13. Når mutasjon rate av en bestemt genotype endringer med miljøet, er det beskrevet som mutasjon-rate plastisitet11. Plastic mutasjon priser har vært mest grundig adressert for stress-indusert mutagenese (SIM)14. I tillegg, ved hjelp av svingninger analyser, har det nylig blitt vist at tettheten som en befolkning av celler vokser (vanligvis en satsvis kultur ved bæreevne) er nært forbundet med mutasjon priser på tvers av bakterier og encellede Landplantenes. Mutasjon rate per Genova per generasjon avtar i tett populasjoner med så mye som 23-fold10,11. Denne tettheten-assosiert mutasjon rate plastisitet (DAMP) kan avhenge av en quorum-sensing system15 og opptre uavhengig av SIM16.
Her er en detaljert protokoll presentert for svingninger analysen brukes til å studere Escherichia coli stamme K-12 å få motstand mot antibiotika Rifampicin i en glukose minimal mediemiljø. Imidlertid bør denne protokollen sees på som en grunnleggende mal som kan utnyttes til å studere en rekke mikrober ved å endre kultur forholdene og fenotypiske markører for mutasjon. Protokollen har utviklet seg fra oppstarten5,17,18,19,20,21,22,23,24,25,26,27,28,29 gjennom bruk på et bredt spekter av mikrober og selv kreftceller30 og har blitt modifisert for å øke gjennomstrømningen, som var avgjørende for riktig testing miljømessige avhengigheter av mikrobiell mutasjon priser10,11,16. Protokollen som beskrives her dekker ikke all metodisk og analytiske spørsmål av svingningene analysen som allerede har vært godt diskutert i litteraturen, spesielt fitness effekter av resistente mutasjoner31, fenotypiske forsinkelse32, celle død33, og egnetheten av ulike algoritmer tilgjengelig for å anslå mutasjon priser26,34. Dette kan være viktig, for eksempel når den miljømessige avhengigheten av fitness effekter kan gi opphav til feilaktig variasjon i mutasjon rate anslag35. Vi registrerer imidlertid at de analytiske verktøyene vi bruker her kan håndtere variasjonen i mutant kondisjon og celle død. Som adressert i notater og diskusjon, er det også anbefalt at flere fenotypiske markører som er usannsynlig å ha samme miljømessig avhengige fitness effekter bli vurdert. Denne protokollen vil gjøre det mulig for folk å rutinemessig analysen miljømessige avhengigheter av mutasjon priser i mangfoldet av mikrobielle stammer og miljøer. Assaying mutasjoner i ulike miljøer har ennå ikke blitt grundig testet og når befolkningstettheten er vurdert, kan svingninger analyser gi et mer presist anslag for mutasjon rate10. Denne protokollen vil muliggjøre flere svingninger analyser skal utføres, som er nødvendig for å forstå mekanismene underbygger mutasjon priser, som igjen er avgjørende for å forstå evolusjon, kreft, aldring, og resistens resistens.
