Hay un número significativo de compuestos disruptores endocrinos (EDC) que se encuentran entre las sustancias más peligrosas en nuestro medio ambiente. Estos son principalmente compuestos estrogénicos que contaminan el agua de los recursos naturales. La diversidad química de las sustancias pertenecientes al grupo dificulta las pruebas de su presencia, ya que se requieren diferentes métodos analíticos para su detección. Basado en su estructura química es muy difícil determinar si una sustancia es realmente capaz de actuar como un estrógeno. Además, estas sustancias nunca están presentes en forma pura en el medio ambiente, por lo que sus efectos pueden verse afectados por otros compuestos, demasiado1. Este problema se puede resolver mediante métodos de detección de efectos, como el uso de biomonitor/bioindicador organismos que muestran efectos estrogénicos2,3,4,5.
Recientemente, una variedad de línea celular6 y sistemas de prueba basados en levadura2,3 se han desarrollado para detectar efectos estrogénicos. Sin embargo, estos son generalmente sólo capaces de detectar la unión de la sustancia al receptor de estrógeno2,3. Además, son incapaces de modelar procesos fisiológicos complejos en el organismo, o para detectar fases sensibles a las hormonas de las etapas de la vida; por lo tanto, a menudo conducen a resultados falsos.
Se sabe que ciertos genes reaccionan sensiblemente al estrógeno en organismos vivos7. La detección de productos genéticos por métodos de biología molecular también es posible en el nivel de proteína o ARNm8,9, pero por lo general implica sacrificio animal. Las leyes de protección animal se han vuelto más estrictas, y existe una creciente demanda de sistemas de ensayo alternativos que minimicen el número y el sufrimiento de los animales utilizados en experimentos o la sustitución del modelo animal por otro modelo10. Con el descubrimiento de proteínas fluorescentes y la creación de líneas de biomarcadores, las tecnologías transgénicas proporcionan una buena alternativa11. Con estas líneas, la activación de un gen sensible al estrógeno se puede probar in vivo.
Entre los vertebrados, el potencial de los peces en la evaluación del riesgo ambiental es excepcional. Ofrecen muchas ventajas sobre los modelos de mamíferos: al ser organismos acuáticos, son capaces de absorber contaminantes a través de todo su cuerpo, producir un gran número de crías, y algunas de sus especies se caracterizan por un corto tiempo de generación. Su sistema endocrino y procesos fisiológicos muestran grandes similitudes con otros vertebrados e incluso con mamíferos, incluidos los humanos12.
También se conocen varios genes para la detección de efectos estrogénicos en peces. Los más importantes son los receptores de estrógeno aromatasa-b, coriogenina-H, y vitellogenina (vtg)7,13. Recientemente, varias líneas de biosensores productores de estrógenos también se han creado a partir de modelos de peces utilizados en el laboratorio, como de pez cebra (Danio rerio)4,5,14,15,16,17. La principal ventaja del pez cebra en la creación de líneas de biosensor es el cuerpo transparente de los embriones y larvas, ya que la señal de reportero fluorescente puede entonces ser fácilmente estudiada in vivo sin sacrificar al animal10. Además de la protección animal, también es una característica valiosa ya que permite estudiar la reacción del mismo individuo en diferentes momentos del tratamiento18.
Estos experimentos utilizan un reportero de vitellogenin linótez de la línea15. La construcción transgene utilizada para el desarrollo de Tg(vtg1:mCherry) tiene un largo (3,4 kbp) promotor natural vitellogenin-1. El receptor de estrógeno (ER) es una proteína potenciadora activada por ligandos que es un representante de la superfamilia de los receptores de esteroides /nuclear. ER se une a secuencias de ADN específicas llamadas elementos de respuesta de estrógeno (EERE) con alta afinidad y transactiva la expresión génica en respuesta al estradiol y otras sustancias estrogénicas, por lo que cuanto más ERE en el promotor causa una respuesta más fuerte19. Hay 17 sitios ERE en la región promotora de la construcción transgén de Tg(vtg1:mCherry) y se espera que imitan la expresión del gen vtg nativo15. Hay una expresión continua de la señal fluorescente en hembras maduradas sexualmente. Sin embargo, en varones y embriones la expresión en el hígado sólo es visible en el tratamiento con sustancias estrogénicas (Figura 1).
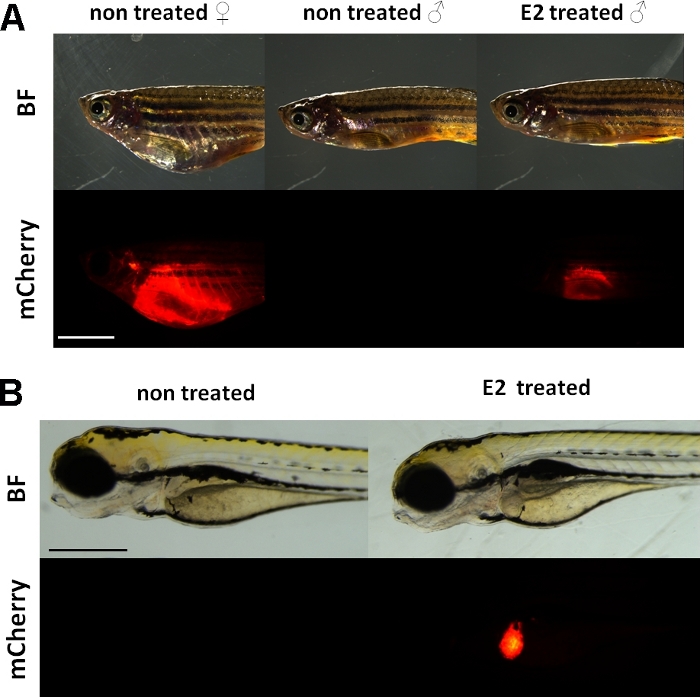
Figura 1: Señal fluorescente roja en el hígado de vg1:mCherry transgenic adult zebrafish y 5 dpf embryos, después de 17-estradiol (E2) inducción. En hembras y en hombres tratados con E2 (25 g/L de tiempo de exposición:48hrs) la fuerte fluorescencia del hígado es visible incluso a través de la piel pigmentada. No se puede ver ninguna señal fluorescente en los machos no tratados (A). Después de la inducción de E2 (50 g/L de tiempo de exposición: 0-120 hpf), también se puede observar una señal fluorescente roja en el hígado de embriones de 5 dpf, que no es visible en los embriones de control (B). Mientras que la señal fluorescente está continuamente presente en las hembras adultas, principalmente los machos y embriones de la línea son adecuados para detectar efectos estrogénicos. (BF: campo brillante, mCherry: vista de filtro fluorescente rojo, imágenes simples, barra de escala A: 5 mm, barra de escala B: 250 m) Haga clic aquí para ver una versión más grande de esta figura.
Al igual que la vitellogenina endógena, el reportero de mCherry sólo se expresa en el hígado. Debido a que la vitellogenina sólo se produce en presencia de estrógeno, no hay señal fluorescente en los controles. Debido a que la expresión está sólo en el hígado, la evaluación de los resultados es mucho más fácil15.
La sensibilidad y usabilidad de los embriones de esta línea se han investigado en diversas mezclas de compuestos estrogénicos y también en muestras ambientales15,20, y en la mayoría de los casos se documentaron las relaciones dosis-respuesta ( Figura2). Sin embargo, en el caso de sustancias altamente tóxicas, principalmente hepatotóxicas ,por ejemplo, zearalenona), sólo una señal fluorescente muy débil puede ser visible en el hígado de embriones tratados y la señal fluorescente de intensidad máxima causada puede alcanzarse dentro de un rango de concentración muy pequeño, lo que dificulta el establecimiento de relaciones dosis-efecto20.
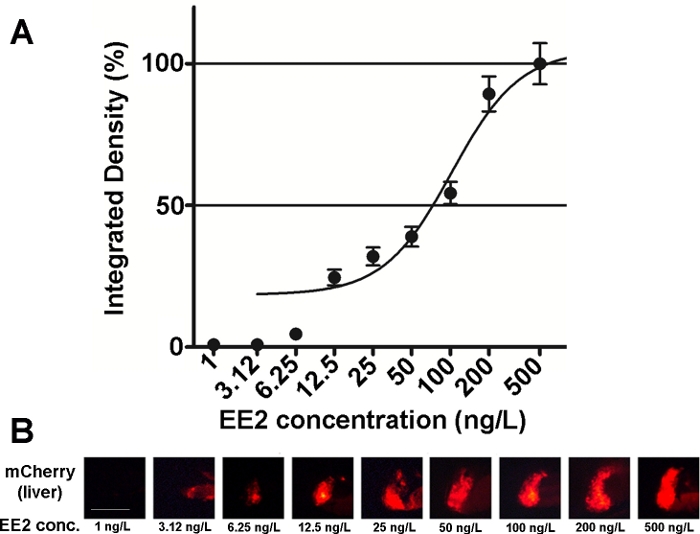
Figura 2: Diagrama de dosis-respuesta (A) e imágenes fluorescentes (mCherry) del hígado (B) expuestos a 17-etilestradiol (EE2), en 5 dpf vtg1:mCherry larvas. Los resultados se expresan como densidad integrada generada a partir de la intensidad de la señal y el tamaño de la zona afectada (-SEM, n a 60). 100% se refiere al máximo observado. La intensidad de la señal fluorescente aumentó gradualmente con la concentración. Barra de escala de 250 m. Haga clic aquí para ver una versión más grande de esta figura.
Hay varias sustancias estrogénicas presentes en el medio ambiente, tales como 17-í-estradiol (concentración ambiental: 0,1–5,1 ng/L)21, 17-etilestradiol (concentración ambiental: 0,16–0,2 g/L)22, zearalenona (concentración ambiental: 0,095–0,22 g/L)23, bisfenol-A (concentración ambiental: 0,45–17,2 mg/L)24. Al probar estas sustancias en forma activa pura con la ayuda de embriones transgénicos mCherry, las concentraciones de efecto observadas más bajas (LOEC) para la detección de signos fluorescentes fueron 100 ng/L para 17-és-estradiol, 1 ng/L para 17-etilestradiol, 100 ng/L para zearalenona y 1 mg/L para el tratamiento con bisfenol-A (96–120 hpf), que está muy cerca o dentro del rango de concentraciones ambientales de las sustancias15. La línea transgénica Tg(vtg1:mCherry) puede ayudar a detectar la estrofica en muestras de aguas residuales después de la exposición directa. La línea es tan sensible como la prueba de estrógeno de levadura de uso común, el ensayo de estrógeno de levadura bioluminiscente (BLYES)15. Con la ayuda de esta línea, se han confirmado los efectos protectores de las beta-ciclodextrinas contra la toxicidad inducida por zearalenona utilizando mezclas químicas20.
En un informe reciente, el uso in vivo de la línea transgénica se demostró con la ayuda de dos metabolitos estrogénicos de zearalenona (ZEA), los anudalenol de los ácidos (ZEA) y los a-zearalenol (a-ZOL y a-ZOL)25. La línea de base del protocolo es apropiada para estudiar los efectos estrogénicos de varios compuestos o muestras ambientales en embriones Tg(vtg1:mCherry).
