Along the physiopathological model from strings to double strings, to small and finally to big clusters, both ECM (Figure 3A) and PCM (Figure 3B) elastic moduli decreased significantly between each pattern change. The only exception was the difference in ECM between strings and double strings (p = 0.072). The results show that the ECM/PCM ratio (Figure 4B) did not change significantly, whereas a marked decrease in the absolute differences in elasticity between ECM and PCM was observed (Figure 4A). Furthermore, the results do not show any significant association concerning the ECM/PCM ratio or associated cellular spatial changes (r = -0.099, p = 0.281).
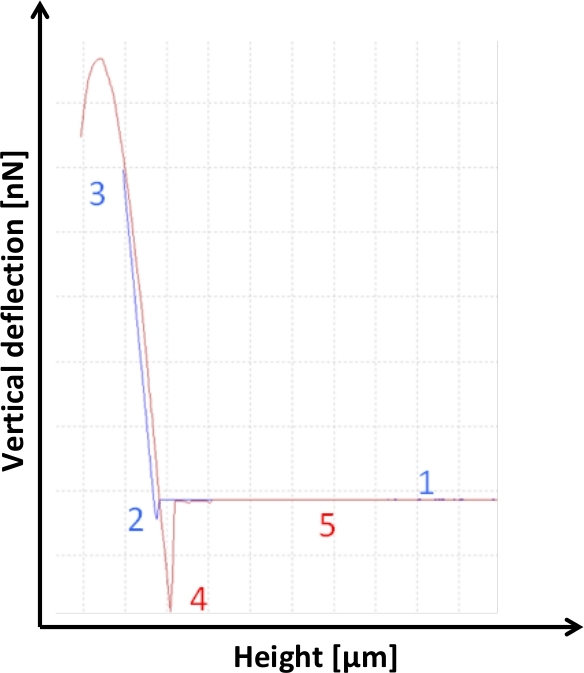
Figure 1: Schematic representation of a force-distance curve in the AFM contact mode. As the probe approaches the surface, the forces are too small to give a measurable deflection of the tip at first, thus leaving the tip in its undisturbed position (1). Then, when the cantilever is very close to the sample, due to adhesive forces active between the tip and the probe, the cantilever actually quickly snaps towards the sample (2). With the probe further approaching the sample, the repulsive deflection then faces against the movement of direction, with an almost linear function of height and deflection until the vertical deflection reaches the relative set point value (3). When retracting (4), in addition to the lowering deflecting forces, adhesion forces are also present while the cantilever is retracted in the Z-axis from the sample. As the AFM probe is pulled off the contact with the sample, it first gets “stuck” before it is able to loosen from the adhesion at the interface, even leading to a short negative deflection of the cantilever, before again reaching its unbent neutral position without contact with the probe (5). The extent of deflection is expressed in the force working on the cantilever expressed in nanonewtons. Please click here to view a larger version of this figure.
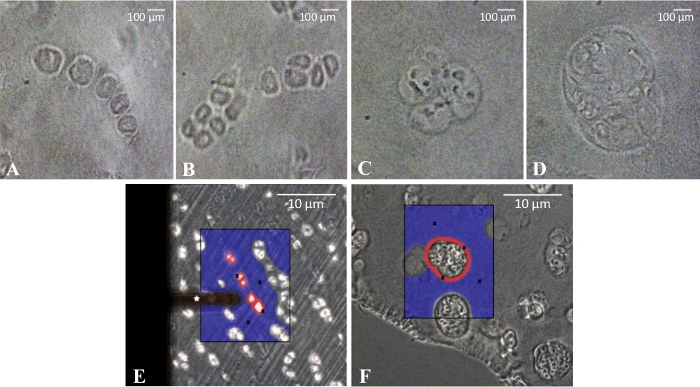
Figure 2: Representative spatial characterization of chondrocytes and AFM measurements of the extracellular matrix (ECM) and pericellular matrix (PCM). (A-D) Characterization of the cellular patterns: strings (A), double strings (B), small clusters (C), and big clusters (D). The elastic moduli of the PCM (red circles) and ECM (blue area) (E/F) were assessed for the different cellular patterns in osteoarthritic cartilage. Measurement sites for the ECM and PCM were selected by the experimenter and are graphically indicated by black crosses. The cantilever tip used for the measurements is marked by a white star. Scale bars represent 10 µm (A-D), and 100 µm (E/F). The figure is adapted and modified from Danalache et al.9. Please click here to view a larger version of this figure.
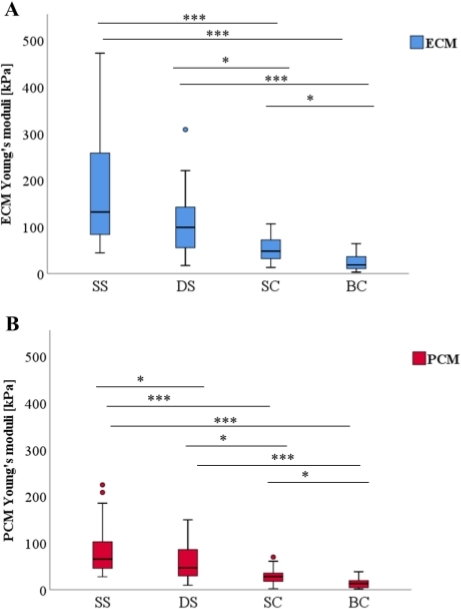
Figure 3: Comparison of the quantified Young’s moduli of the extracellular matrix (ECM) and the pericellular matrix (PCM) as a function of spatial chondrocyte organization. With progressive pathological spatial chondrocyte organization, a gradual decrease of elasticity was noted in the boxplots for both the ECM (A) and the PCM (B) (*p < 0.05, ***p < 0.001). Abbreviations: SS: single strings, DS: double strings, SC: small clusters, BC: big clusters. The figures are taken from Danalache et al.9. Please click here to view a larger version of this figure.
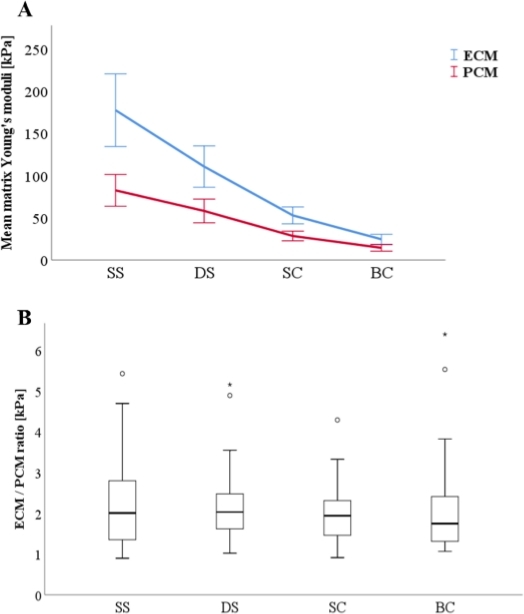
Figure 4: Relationship of the Young’s moduli of the extracellular matrix (ECM) and the pericellular matrix (PCM) as a function of cellular spatial organization. An increasingly pathological spatial chondrocyte organization was associated with a decrease of the Young’s moduli for both ECM and PCM (A). While these spatial changes took place, the ratio of ECM and PCM elasticity remained constant, showing no significant changes (B). The data are presented as a line diagram with mean ± standard error (A) and boxplots (B). Abbreviations: SS: single strings, DS: double strings, SC: small clusters, BC: big clusters. The figures are taken from Danalache et al.9. Please click here to view a larger version of this figure.
Supplemental Figure 1: Representative force curve obtained by indentation of the pericellular matrix (PCM) of a single string pattern showing the fit results (orange arrow) as well as the residual root mean square (residual RMS; black arrow). The fit results include the contact point between sample and tip, the Young’s Modulus, and the baseline. The residual RMS displayed below describes the difference between the fit and the force data, thereby representing the quality of a force curve fit. Please click here to download this figure.
| Table 1. Parameters for gluing a microsphere on the AFM-probe | |
| Parameters | Value |
| Setpoint | 5.0 V |
| Adjust baseline | 1 |
| Pulling length | 90.0 µm |
| Z movement | Constant speed |
| Extend speed | 5.0 µm/s |
| Extend time | 18.0 s |
| Contact time | 90.0 s |
| Delay mode | Constant force |
| Sample rate | 2000 Hz |
Table 1. Parameters for gluing a microsphere on the AFM-probe.
| Table 2. Approach parameters | |
| Approach Parameters | Value |
| Approach IGain | 5.0 Hz |
| Approach PGain | 0.0002 |
| Approach target height | 10.0 µm |
| Approach setpoint | 5.00 V |
| Approach baseline | 0.00 V |
Table 2. Approach parameters.
| Table 3. Run parameters | |
| Parameters | Value |
| Setpoint | 1.0 V |
| Adjust baseline | 1 |
| Pulling length | 90.0 µm |
| Z movement | Constant speed |
| Extend speed | 5.0 µm/s |
| Extend time | 18.0 s |
| Contact time | 0.0 s |
| Delay mode | Constant force |
| Sample rate | 2000 Hz |
Table 3. Run parameters.
