人类细胞不断暴露在各种来源的DNA破坏剂中。外源主要包括接触辐射、化学品(包括化疗剂和某些抗生素)和病毒,而主要的内源包括DNA复制和氧化应激方面的错误。基因毒性暴露的直接影响范围从改良的碱基到潜在的致命DNA双链断裂(DSB),这取决于压力和暴露剂量。最终,未修复或修复不当的DNA损伤可导致突变的积累、基因组重组、基因组不稳定,并最终导致致癌1。哺乳动物细胞已经进化出复杂的途径来识别特定类型的DNA损伤2,2,3,并及时修复它们,与细胞周期进展同步。
电离辐射(IR)损害DNA双螺旋,并造成双链断裂(DSB),这是最有害的DNA损伤形式之一。MRN (MRE11, RAD50, NBS1) 复合功能作为 DNA 末端的传感器,并激活蛋白激酶 ataxia telangiectasia 突变 (ATM)4,5。在DNA端对ATM的初始激活后,ATM在断点触发一系列DDR事件,启动一个关键事件,即组蛋白变种H2AX6的磷酸化。在残留物S139上的H2AX磷酸化激活它到+H2AX,跨越区域,高达围绕DNA病变6,7,8,97,8的巨型碱。6这一事件增加了DNA的可访问性,导致其他DNA修复蛋白的招募和积累7。由于 +H2AX 是丰富和具体诱导周围的 DSB,它可以很容易地可视化使用特定的抗体,并通常用作 DSB 在 DNA 修复领域的代理标记。一旦中断被发出信号,细胞激活其DNA修复途径并处理DNA损伤。蛋白质MDC1(DNA损伤检查点蛋白1的中介)直接结合+H2AX10,与ATM11和NBS1 12,13,相互作用。它有助于提高 MRN 复合物在 DSB 的浓度,并启动正 ATM 反馈回路。•一旦修复断裂,快速清除H2AX,从而允许监控DSB间隙。其次是显微镜,随着时间的推移,μH2AX 的减少提供了残余断裂和 DNA 修复效率的间接测量。
真核细胞可以通过多种途径修复DSB,两个主要途径是非同源端连接(NHEJ)和同源重组(HR)(图1)。NHEJ基本上连接DNA双链结束不使用扩展同源和在整个细胞周期14,15运行。14,HR在S和G2阶段成为占主导地位,否则被压抑,因为它需要一个姐妹色度作为一个同源模板修复14,16。14,NHEJ和HR之间的路径选择不仅取决于姐妹色度的物理接近性,还取决于DNA端切除17的延伸,它抑制了NHEJ。
同源性依赖的DSB修复启动核解降解的5’链从断裂端产生3’单链DNA(ssDNA)尾部,这个过程被称为5′-3’切除。MRN复合物启动DNA端切除术,并结合BLM/EXO1(布洛姆综合征蛋白/外核酶1)或BLM/DNA2(DNA复制ATP依赖性直升机酸酶,/核酸酶)18、19、20、21、22,19,20进行进一,步切除。21CtIP(CtBP相互作用蛋白)通过与 MRN 复合物23的直接相互作用和 BRCA1(乳腺癌 1 型易感性蛋白)24,25的招募来增强 DNA 末切。复制蛋白A(RPA)迅速结合到暴露的ssDNA,然后被重组酶蛋白RAD51取代,形成核蛋白丝,催化同源搜索和链入侵26,27,28。26,27,28
切除的启动是修复路径选择的关键步骤。一旦切除开始,DNA末端成为由Ku70/Ku80异质剂(NHEJ通路的组成部分)结合的不良基质,细胞被承诺到HR 17,29,30。17,29,30Ku70/Ku80异质剂结合到DSB末端,招募DNA-PKcs和p53结合蛋白,1(53BP1)29,2930。53BP1在G1中起到切除屏障的作用,从而在推广NHEJ31、32,32的同时阻断HR,但在S相中以BRCA1依赖的方式去除,从而允许切除发生33,34,34。因此,53BP1 和 BRCA1 在 DSB 维修中扮演相反的角色,53BP1 是 NHEJ 促进者,而 BRCA1 则通过 HR 进行中断修复。
在实验室中,DSB形成可由电离辐射(IR)引起。虽然本示例利用高剂量的 4 Gy、1 Gy 和 2 Gy 也创建大量的 DSB,适合通过丰富的蛋白质分析叶层形成。需要注意的是,使用的辐射类型和剂量可导致DNA和细胞中不同的病变:虽然IR诱导DSB,它也可能导致单股断裂或碱基修饰(见35,36,,36关于辐照线性能量转移(LET)和DNA损伤类型的参考)。为了确定电离辐射诱导的源(IRIF)形成的动力学及其间隙,表明修复激活的DDR8、9、37、38,9,37,的损伤和反转,在电离辐射后的不同时间点可以监测源层。所有主要DNA损伤蛋白的激活和清除时间已知为39种,许多被调查为关键事件的代理标记。例如,对 ssDNA 具有高亲和力的 pRPA 用作断分切除的代理,MRN 蛋白(MRE11、RAD50、NBS1)和外核酶也可用于评估切除效率。虽然RAD51、BRCA1、BRCA2(乳腺癌2型易感性蛋白)和PALB2(BRCA2的合作伙伴和本地化者)可以监测,以评估人力资源效率,而Ku蛋白或53BP1的存在则用作NHEJ的标记(图1)。
当DNA修复机械的蛋白质相互招募到超级复合物中分离和组装时,DNA蛋白和蛋白质-蛋白质的相互作用可以通过跟踪它们各自的定位并分析蛋白质的共同定位来推断,如细胞40、41、42,41,42中的重叠信号。在细胞系中,通过基因组编辑或对质粒突变体过度表达,在特定DNA修复基因中引入点突变或删除,允许调查特定残留物及其在识别DNA损伤(例如,与+H2AX)或复杂组装(与其他蛋白质或几种蛋白质共同定位)中可能发挥的作用,以及它们对DNA修复的影响。在这里,我们使用间接免疫荧光作为一种均值,通过跟踪 +2AX foci 来研究 DSB 的形成和分辨率。我们还提出了一个示例,由 DSB 修复的主要参与者进行 foci 形成和共同定位分析:p53 结合蛋白 1 (53BP1)32。如前所述,53BP1 被认为是 DNA 修复路径选择的核心。在 53BP1 积累及其与 +2AX 的共定位后,提供了有关细胞周期相、DNA损伤积累和用于修复 DSB 的路径的宝贵信息。间接免疫定位的目的是评估细胞系DNA损伤修复的效率,如本研究中的IR之后,或在接触细胞中的各种应力后,从DNA交联到复制分叉的阻塞(表1提供了DNA损伤剂列表)。
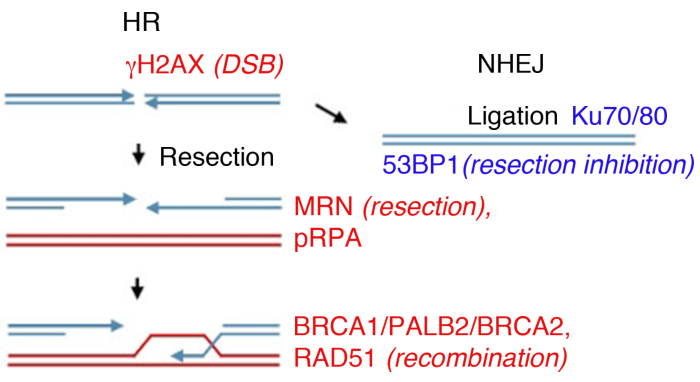
图1:DNA双链断裂(DSB)修复途径。
DSB 修复涉及两个主要途径:同源重组(HR、左)和非同源端连接(NHEJ,右)。断裂后,蛋白质被激活以标记断裂(+2AX),参与最终切除(MRN),涂上被切除的ssDNA(pRPA),促进重组(BRCA1,PALB2,BRCA2,RAD51)或限制切除,促进NHEJ(53BP1)。其他蛋白质参与损伤修复,但列出的蛋白质通常遵循间接免疫荧光。 请单击此处查看此图的较大版本。
| DNA损伤剂 | 行动机制 | 推荐剂量 |
| • 射线/X 射线 | 辐射 形成双链断裂与一些不受控制的细胞效应 |
1-4 Gy |
| 36Ar 离子 | 辐射 双链断裂的形成 |
270千瓦/μm |
| • 粒子 | 辐射 双链断裂的形成 |
116 keV/μm |
| 博莱 霉 素 | DNA合成的抑制剂 | 0.4-2 μg/mL |
| 喜 树 碱 | 托托莫默酶 I 的抑制剂 | 10-200 nM |
| 顺 铂 | 阿尔基拉特剂 (诱导海峡内部交联) |
0.25-2 μM |
| 阿 霉 素 | 中间代理 托托莫默酶II.的抑制剂 |
10-200 nM |
| 埃托波赛德 | 托托莫默酶II.的抑制剂 | 10 μM |
| 羟基尤雷亚 | DNA合成的抑制剂 (由核糖核酸还原酶) |
10-200 μM |
| 甲基甲烷硫化甲酸酯 | 阿尔基拉特剂 | 0.25-2 mM |
| 米托米霉素 C | 阿尔基拉特剂 | 0.25-2 μM |
| 紫外线(UV)光 | 甲状腺素二丁的形成 (产生DNA链失真) |
50-100 mJ/厘米2 |
表1:基因毒性剂.DNA损伤剂的例子,其作用机制和根据建议的工作浓度引起的损伤。
