Menneskelige celler er konstant udsat for en række DNA skadelige stoffer af forskellig oprindelse. Eksogene kilder består for det meste af eksponering for stråling, kemikalier (herunder kemoterapeutiske midler og visse antibiotika) og virus, mens de vigtigste endogene kilder omfatter fejl i DNA-replikation og oxidativt stress. De direkte virkninger af genotoksisk eksponering kan variere fra en modificeret base til en potentielt dødelig DNA dobbelt-streng pause (DSB), afhængigt af stress og eksponeringsdosis. I sidste ende, urepareret eller fejlrepareret DNA-skader kan føre til ophobning af mutationer, genomiske rearrangements, genom ustabilitet og i sidste ende føre til carcinogenese1. Pattedyrceller har udviklet komplekse veje til at genkende specifikke typer afDNA-skader 2,,3 og reparere dem i tide, synkroniseret med cellecyklus progression.
Iioniserende stråling (IR) skader DNA dobbelt helix og skaber dobbelt-streng pauser (DSBs), en af de mest skadelige former for DNA-skader. MRN (MRE11, RAD50, NBS1) kompleks fungerer som en sensor af DNA ender og aktiverer protein kinase ataxia telangiectasia muteret (ATM)4,5. Efter den første aktivering af ATM ved DNA ender, ATM udløser en kaskade af DDR begivenheder på det sted, hvor pausen, indlede med en vigtig begivenhed, fosforylering af histone variant H2AX6. H2AX-fosforylering på restprodukt S139 aktiverer det i γH2AX, der spænder over områder op til megabaser omkring DNA-læsionen6,7,8,9. Denne hændelse øger DNA-tilgængeligheden, hvilket fører til rekruttering og akkumulering af andre DNA-reparationsproteiner7. Fordi γH2AX er rigeligt og specifikt induceret omkringliggende DSBs, det kan let visualiseres ved hjælp af specifikke antistoffer, og er almindeligt anvendt som en surrogat markør for DSBs i DNA reparation området. Når bruddet er signaleret, celler aktivere deres DNA reparation veje og behandle DNA-skader. Proteinet MDC1 (mægler af DNA-skader checkpoint protein 1) binder direkte γH2AX10, interagerer med ATM11 og også med NBS112,13. Det bidrager til at øge koncentrationen af MRN-komplekset i DSB og indleder et positivt atm-feedback-loop. γH2AX fjernes hurtigt, når bruddet er repareret, hvilket gør det muligt at overvåge DSB-frigangen. Efterfulgt af mikroskopi giver faldet i γH2AX over tid en indirekte måling af resterende brud og DNA-reparationseffektivitet.
Eukaryote celler kan reparere DSBs ved flere veje, de to vigtigste er ikke-homologe end-sammenføjning (NHEJ) og homolog rekombination (HR) (Figur 1). NHEJ i det væsentlige ligaerer DNA dobbelt-streng ender uden brug af udvidet homologi og opererer i hele cellecyklus 14,15. HR bliver fremherskende i S- og G2-faser, og undertrykkes ellers, da det kræver en søsterkromet som en homolog skabelon til reparation14,16. Pathway valg mellem NHEJ og HR ikke kun afhænger af den fysiske nærhed af søsterchromatid, men også af udvidelsen af DNA ende resektion17, som hæmmer NHEJ.
Homologi-afhængige DSB reparation initiere ved nukleolytisk nedbrydning af 5 ‘streng fra pausen ender til at generere 3 ‘single-streng DNA (ssDNA) haler, en proces benævnt 5’-3 ‘resektion. MRN-komplekset indleder DNA-slutresektion, og yderligere resektion behandles i kombination med BLM/EXO1 (Bloom syndrom protein/exonuclease 1) eller BLM/DNA2 (DNA replikation ATP-afhængig helicase/nuklease)18,19,20,21,22. DNA-slutresektion forstærkes af CtIP (CtBP-interagerende protein) gennem dets direkte interaktion med MRN-kompleks23 og rekruttering af BRCA1 (brystkræfttype 1 modtagelighedsprotein)24,25. Replikering protein A (RPA) straks binder sig til ssDNA eksponeret og derefter fordrevet af recombinase protein RAD51 til at danne en nukleoprotein glødetråd, der katalyserer homolog søgning og strand invasion26,27,28.
Indledningen af resektion er et kritisk skridt for reparation pathway valg. Når resektion er indledt, DNA ender bliver fattige substrater til binding af Ku70/Ku80 heterodimer (komponent af NHEJ vej) og celler er forpligtet til HR17,,29,30. Den Ku70/Ku80 heterodimer binder sig til DSB ender, rekruttere DNA-pc’er og p53 Bindende Protein 1 (53BP1)29,30. 53BP1 fungerer som en hindring for resektion i G1 og blokerer således HR og fremmer samtidig NHEJ31,32, men fjernes på en BRCA1-afhængig måde i S-fasen , hvilket gør det muligt at genskrive33,34. Derfor spiller 53BP1 og BRCA1 modsatrettede roller i DSB-reparationen, hvor 53BP1 er NHEJ-facilitator, mens BRCA1 handler, der gør det muligt at reparere gennem HR.
I laboratoriet kan DSB-formationen induceres ved iioniseringsstråling (IR). Mens dette eksempel udnytter en høj dosis på 4 Gy, 1 Gy og 2 Gy også skabe en betydelig mængde af DSBs, egnet til analyse af foci dannelse af rigelige proteiner. Det er vigtigt at bemærke, at den anvendte type og dosis af stråling kan føre til forskellige læsioner i DNA og i cellen: mens IR inducerer DSBs, kan det også forårsage enkelt streng pauser eller base modifikation (se35,36 for en reference på bestråling lineær energioverførsel (LET) og type DNA-skader). For at bestemme kinetik af iionizing stråling-induceret foci (IRIF) dannelse og deres clearance, som indikerer reparation af skader og tilbageførsel af den aktiverede DDR8,9,37,38, foci dannelse kan overvåges på forskellige tidspunkter efter iionizing stråling.9 Timing af aktivering og clearance af alle større DNA-skader proteiner er kendt39, og mange er undersøgt som surrogat markører for vigtige begivenheder. For eksempel, pRPA, som har høj affinitet for ssDNA bruges som et surrogat for pause resektion, MRN proteiner (MRE11, RAD50, NBS1) og exonucleaser kan bruges til at vurdere resektion effektivitet også. Mens RAD51, BRCA1, BRCA2 (brystkræft type 2 modtagelighed protein), og PALB2 (partner og localizer af BRCA2) kan overvåges for at evaluere HR effektivitet, tilstedeværelsen af Ku proteiner eller 53BP1, anvendes som markører for NHEJ (Figur 1).
Som proteiner af DNA reparation maskiner rekruttere hinanden til pause og samle i super-komplekser, DNA-protein og protein-protein interaktioner kan udledes ved at følge deres individuelle lokalisering over tid og analysere co-lokalisering af proteiner, som visualiseret af overlappende signaler i celle40,41,42. I cellelinjer giver indførelsen af punktmutationer eller sletning i specifikke DNA-reparationsgener enten gennem genomredigering eller ved overekspression af plasmidbaserede mutanter mulighed for undersøgelse af specifikke restkoncentrationer og deres mulige rolle i genkendelse af DNA-skader (f.eks. co-lokalisering med γH2AX) eller kompleks samling (sam lokalisering med en anden eller flere proteiner) samt deres indvirkning på DNA-reparationen. Her bruger vi indirekte immunfluorescens som et middel til at undersøge dannelsen og opløsningen af DSB’er ved at følge γH2AX foci over tid. Vi præsenterer også et eksempel på foci formation og co-lokalisering analyse af en stor aktør i DSB reparation: p53 Bindende Protein 1 (53BP1)32. Som tidligere nævnt, 53BP1 anses for centralt for DNA reparation pathway valg. Efter 53BP1 ophobning og dens co-lokalisering med γH2AX giver værdifulde oplysninger om celle cyklus fase, DNA-skader ophobning, og vej bruges til at reparere DSBs. Formålet med indirekte immunolokalkisering er at vurdere effektiviteten af DNA-skade reparation i cellelinjer, efter IR som i denne undersøgelse, eller efter eksponering for forskellige belastninger i celle, fra DNA crosslinking til blokering af replikation gaffel (en liste over DNA skadelige stoffer er fastsat i tabel 1).
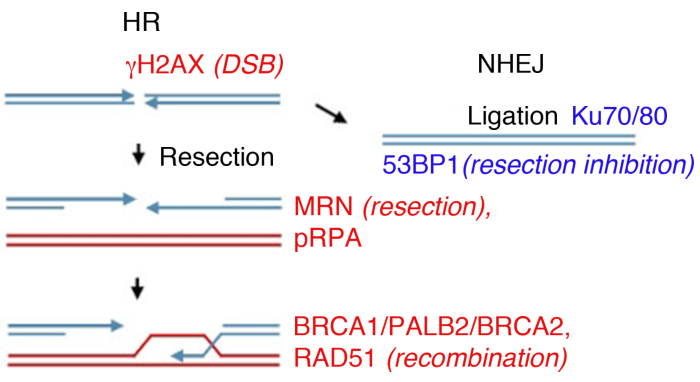
Figur 1: DNA dobbelt streng pauser (DSB) reparation veje.
DSB reparation indebærer to store veje: Homolog rekombination (HR, venstre) og ikke-homologe End-Joining (NHEJ, højre). Efter pausen aktiveres proteiner for at markere bruddet (γH2AX), deltager i slutresektionen (MRN), belægger den resected ssDNA (pRPA), fremmer rekombination (BRCA1, PALB2, BRCA2, RAD51) eller begrænser resektion og fremmer NHEJ (53BP1). Andre proteiner deltager i skadesreparation, men de anførte proteiner efterfølges rutinemæssigt af indirekte immunfluorescens. Klik her for at se en større version af dette tal.
| DNA-skademiddel | Virkningsmekanisme | Anbefalet dosis |
| γ-stråler/røntgenstråler | Stråling Dannelse af dobbeltstrengede brud med nogle ukontrollerede cellulære effekter |
1-4 Gy |
| kr. Ar ioner | Stråling Dannelse af dobbeltstrengede pauser |
270 keV/μm |
| α-partikler | Stråling Dannelse af dobbeltstrengede pauser |
116 keV/μm |
| Bleomycin | Hæmmer af DNA-syntese | 0,4-2 μg/ml |
| Camptothecin | Hæmmer af topoisomerase I | 10-200 nM |
| Cisplatin | Alkylerende middel (inducerende intrastrand krydslinks) |
0,25-2 μM |
| Doxorubicin | Intercaleringsmiddel Hæmmer af topoisomerase II |
10-200 nM |
| Etoposide | Hæmmer af topoisomerase II | 10 μM |
| Hydroxyurea | Hæmmer af DNA-syntese (ved ribonucleotid reduktase) |
10-200 μM |
| Methylmethanulfonat | Alkylerende middel | 0,25-2 mM |
| Mitomycin C | Alkylerende middel | 0,25-2 μM |
| Ultraviolet (UV) lys | Dannelse af thymidindæmpere (genererer forvrængning af DNA-kæden) |
50-100 mJ/cm2 |
Tabel 1: Genotoksiske midler. Eksempler på DNA-skadelige stoffer, deres virkningsmekanisme og de skader, der er forårsaget på grundlag af foreslået arbejdskoncentration.
