תאים אנושיים נחשפים כל הזמן למגוון של סוכני DNA מזיקים ממקורות שונים. מקורות אקסוגניים מורכבים בעיקר מחשיפה לקרינה, כימיקלים (כולל חומרים כימותרפיים וכמה אנטיביוטיקה), ווירוסים, בעוד המקורות האנדורניים העיקריים כוללים שגיאות בשכפול DNA ומתח חמצוני. ההשפעות הישירות של חשיפה גנוטוקסית יכול לנוע מבסיס שונה לשבור DNA קטלני פוטנציאלית דו גדיל (DSB), בהתאם ללחץ ואת מינון החשיפה. בסופו של דבר, נזק DNA לא מתוקן או שלא תוקן כהלכה יכול להוביל להצטברות של מוטציות, סידורים מחדש גנומיים, חוסר יציבות הגנום ובסופו של דבר להוביל קרסינוגנזה1. תאים יונקים פיתחו מסלולים מורכבים כדי לזהות סוגים ספציפיים של נזק DNA2,,3 ולתקן אותם בזמן, מסונכרן עם התקדמות מחזור התא.
קרינה מיננת (IR) פוגעת בסליל ה-DNA הכפול ויוצרת הפסקות דו-גדיליות (DSBs), אחת הצורות המבזות ביותר של נזק לדנ”א. MRN (MRE11, RAD50, NBS1) פונקציות מורכבות כמו חיישן של ה-DNA מסתיים ומפעיל את החלבון קינאז ataxia telangiectasia מוטציה (ATM)4,,5. לאחר ההפעלה הראשונית של ATM על ידי קצוות DNA, ATM מפעיל מפל של אירועי DDR באתר ההפסקה, ייזום עם אירוע מפתח, phosphorylation של משתנה histone H2AX6. H2AX phosphorylation על שאריות S139 מפעיל אותו לתוך ơH2AX, משתרע על פני אזורים עד megabases סביב נגע ה-DNA6,7,,8,9. אירוע זה מגביר את הנגישות DNA, המוביל גיוס והצטברות של חלבוני תיקון DNA אחרים7. מכיוון 8H2AX הוא בשפע ובמיוחד מושרה סביב DSBs, זה יכול להיות חזותי בקלות באמצעות נוגדנים ספציפיים, והוא משמש בדרך כלל כסמן פונדקאית עבור DSBs בשדה תיקון ה-DNA. ברגע שההפסקה מאותתת, תאים מפעילים את נתיבי תיקון הדנ”א שלהם ומעבדים את נזקי הדנ”א. החלבון MDC1 (מגשר של חלבון מחסום נזק DNA 1) ישירות נקשר ơH2AX10, אינטראקציה עם ATM11 וגם עם NBS112,13. זה תורם להגדלת הריכוז של מתחם MRN ב DSB וייזום לולאת משוב חיובית כספומט. ơH2AX מוסר במהירות לאחר תיקון השבר, כתוצאה מכך, המאפשר ניטור של סיווג DSB. ואחריו מיקרוסקופיה, הירידה ב 0H2AX לאורך זמן מספק מדידה עקיפה של הפסקות שיורית ויעילות תיקון DNA.
תאים אאוקריוטיים יכולים לתקן DSBs על ידי מספר מסלולים, שני העיקריים להיות לא הומולוגי סוף הצטרפות (NHEJ) ו שילוב הומולוגי (HR)(איור 1). NHEJ למעשה רצועות DNA כפול גדיל מסתיים ללא שימוש הומולוגיה מורחבת ופועללאורך כל מחזור התא 14,15. משאבי אנוש הופכים לדומיננטיים בשלבי S ו-G2, ומודחקים באופן אחר, שכן היא דורשת כרומטיד אחות כתבנית הומולוגיתלתיקון 14,16. בחירת מסלול בין NHEJ ו HR לא רק תלוי הקרבה הפיזית של כרומט אחותו, אלא גם על הרחבת ה-DNAסוף חותך 17, אשר מעכב NHEJ.
תיקון DSB תלוי הומולוגיה יוזם על ידי השפלה נוקליאוליטית של 5 ‘ גדיל מן קצוות ההפסקה כדי ליצור 3 ‘ זנבות DNA גדיל יחיד (ssDNA), תהליך המכונה 5′-3’ ריתוק. תסביך MRN יוזם את החיתוך קצה DNA וכתור נוסף מעובד בשילוב עם BLM / EXO1 (תסמונת בלום חלבון / exonuclease 1) או BLM / DNA2 (שכפול DNA ATP תלוי הליקאז / nuclease)18, 19,,20,21,22.19 ניתוח קצה DNA משופרת על ידי CtIP (חלבון אינטראקציה CtBP) באמצעות האינטראקציה הישירה שלה עם MRNמורכב 23 וגיוס של BRCA1 (סרטן השד סוג 1 חלבון רגישות)24,25. חלבון שכפול A (RPA) נקשר מיד ssDNA חשוף ולאחר מכן נעקר על ידי חלבון רקומבינאז RAD51 כדי ליצור חוט נוקליאופרוטאין כי מזרז חיפוש הומולוגיופלישה גדיל 26,27,28.
החניכה של ניתוח הוא צעד קריטי לבחירה במסלול תיקון. לאחר ההתקנה, מסתיים ה-DNA להיות מצעים עניים עבור איגוד על ידי Ku70/Ku80 הטרודימר (רכיב של מסלול NHEJ) ותאים מחויבים HR17,29,30. Ku70/Ku80 הטרודימר נקשר לקצוות DSB, גיוס DNA-PKcs ו p53 מחייב חלבון 1 (53BP1)29,,30. 53BP1 משמש כמחסום לניתוק ב-G1, ובכך חוסם את משאבי האנוש תוך קידום NHEJ31,32, אך הוא מוסר באופן תלוי BRCA1 בשלב S, וכתוצאה מכך מאפשר לניתוחפעולה 33,,34. לכן, 53BP1 ו BRCA1 לשחק תפקידים מנוגדים בתיקון DSB, עם 53BP1 להיות מנחה NHEJ בעוד פעולות BRCA1 המאפשר הפסקות לתקן באמצעות משאבי אנוש.
במעבדה, היווצרות DSB יכול להיות מושרה על ידי קרינה מיונן (IR). בעוד דוגמה זו מנצלת מינון גבוה של 4 Gy, 1 Gy ו 2 Gy גם ליצור כמות משמעותית של DSBs, מתאים לניתוח של היווצרות foci על ידי חלבונים בשפע. חשוב לציין כי הסוג והמינון של קרינה בשימוש יכול להוביל נגעים שונים ב-DNA ובתא: בעוד IR גורם DSBs, זה יכול גם לגרום הפסקות גדיל יחיד או שינוי בסיס(ראה 35,36 לעיון על העברת אנרגיה ליניארית קרינה (LET) וסוג של נזק DNA). כדי לקבוע את הקינטיקה של היווצרות foci (IRIF) הנגרמת על-ידי קרינה מיוננת ואת הסיווג שלהם, אשר מצביעים על תיקון של הנזק והיפוך של DDRמופעל 8,9,37,38,היווצרותfoci ניתן לפקח בנקודות זמן שונות לאחר קרינה מיוננת. תזמון ההפעלה והאישור של כל החלבונים העיקריים לנזקי DNAידוע 39, ורבים נחקרים כסמן חלופי של אירועי מפתח. לדוגמה, pRPA, אשר בעל זיקה גבוהה עבור ssDNA משמש כתחליף של ניתוק לשבור, חלבוני MRN (MRE11, RAD50, NBS1) ו exonucleases יכול לשמש כדי להעריך יעילות השבירה מדי. בעוד RAD51, BRCA1, BRCA2 (סרטן השד סוג 2 חלבון רגישים), ו PALB2 (שותף ו localizer של BRCA2) ניתן לפקח כדי להעריך את יעילות משאבי אנוש, הנוכחות של חלבוני Ku או 53BP1, משמשים סמנים של NHEJ (איור 1).
כמו חלבונים של מכונות תיקון DNA לגייס אחד את השני לשבור ולהרכיב בסופר קומפלקסים, אינטראקציות DNA-חלבון וחלבון-חלבון ניתן להסיק על ידי ביצוע מקומיות הפרט שלהם לאורך זמן וניתוח התאמה חברתית של חלבונים, כפי שניתן לראות על ידי אותותחופפים בתא 40,41,42. בקווי תאים, המבוא של מוטציות נקודתיות או מחיקה בגנים ספציפיים לתיקון DNA או באמצעות עריכת הגנום, או על ידי דיכוי יתר של מוטציות מבוססות פלסמיד, מאפשר חקירה של שאריות ספציפיות ותפקידם האפשרי בהכרה בנזקי DNA (למשל, התאמה משותפת עם ΩH2AX) או הרכבה מורכבת (התאמה משותפת עם אחר, או מספר, חלבונים), כמו גם השפעתם על תיקון DNA. כאן, אנו משתמשים בחיסונים עקיף כדרך לחקור את היווצרות ורזולוציה של DSBs על ידי ביצוע 8H2AX foci לאורך זמן. אנו מציגים גם דוגמה אחת של היווצרות foci וניתוח התאמה למיקום אחר-תום-מקומי על-ידי שחקן מרכזי בתיקון DSB: p53 Binding Protein 1 (53BP1)32. כפי שהוזכר קודם לכן, 53BP1 נחשב מרכזי לבחירה מסלול תיקון DNA. לאחר הצטברות 53BP1 והתמצאות התקצוב ההפוך שלה עם ơH2AX מספק מידע יקר על שלב מחזור התא, הצטברות נזק DNA, מסלול המשמש לתיקון DSBs. המטרה של אימונוקולציה עקיפה היא להעריך את היעילות של תיקון נזק DNA בקווי תאים, בעקבות IR כמו במחקר זה, או לאחר חשיפה ללחצים שונים בתא, מן הצלבת DNA לחסימה של שקע שכפול (רשימה של סוכנים מזיקים DNA מסופק בטבלה 1).
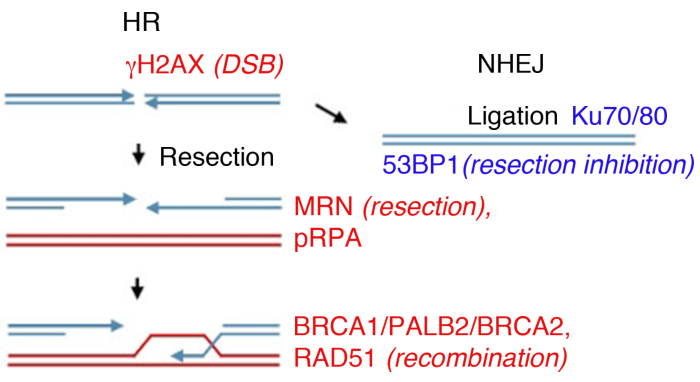
איור 1: נתיבי תיקון של שוברי גדילים כפולים של דנ”א (DSB).
תיקון DSB כולל שני מסלולים עיקריים: שילוב מחדש הומולוגי (HR, שמאל) והצטרפות לסוף לא הומולוגית (NHEJ, נכון). לאחר ההפסקה, חלבונים מופעלים כדי לסמן את ההפסקה (ơH2AX), להשתתף כריתה סוף (MRN), לכסות את ssDNA החתוכים (pRPA), לקדם שילוב מחדש (BRCA1, PALB2, BRCA2, RAD51) או להגביל את כריתת ולקדם NHEJ (53BP1). חלבונים אחרים להשתתף בתיקון נזק, אבל חלבונים המפורטים באופן שגרתי מלווה אימונו-לודיזם עקיף. לחץ כאן כדי להציג גירסה גדולה יותר של נתון זה.
| סוכן מזיק DNA | מנגנון פעולה | מינון מומלץ |
| ơ-קרני רנטגן/ קרני רנטגן | קרינה היווצרות של הפסקות דו-גדיליות עם השפעות תאיות בלתי מבוקרות |
1-4 ג’י |
| 36 00:00: יונים | קרינה היווצרות של הפסקות דו-גדיליות |
270 קילוו/μm |
| α-חלקיקים | קרינה היווצרות של הפסקות דו-גדיליות |
116 keV/μm |
| בלומיצין (Bleomycin) | מעכב סינתזת דנ”א | 0.4-2 μg/mL |
| (קמפטוסין, שוויץ) | מעכב של טופואיסומראז אני | 10-200 00 00 00 00:00:00,00 |
| צ’יספלטין (1999) | סוכן אלקילטינג (גורם קישורים תוך סטריים והצלבה) |
0.25-2 μM |
| דוקסורביצין (11 | סוכן התבות מעכב של טופואיסומראז II |
10-200 00 00 00 00:00:00,00 |
| אטופוסייד (אטופוסייד) | מעכב של טופואיסומראז II | 10 μM |
| הידרוקסיורה (הידרוקסיורה) | מעכב סינתזת דנ”א (על ידי רדוקטסה ריבונוקלאוטיד) |
10-200 μM |
| מתיל מתנסולפונט | סוכן אלקילטינג | 0.25-2 מ”מ |
| מיטומיצין C | סוכן אלקילטינג | 0.25-2 μM |
| אור אולטרה סגול (UV) | היווצרות דימי דימידין (יצירת עיוות של שרשרת ה-DNA) |
50-100 מ”ג/ס”מ2 |
שולחן 1: סוכנים גנוטוקסיים. דוגמאות של סוכני DNA מזיקים, מנגנון הפעולה שלהם ואת הנזק המושרה בהתבסס על ריכוז עבודה המוצע.
