Application of DH31 against unstimulated MTs results in a significant increase in fluid secretion rate, confirming its role as a diuretic hormone in Aedes mosquitoes (Figure 1A). When tubules are treated with AedaeCAPA-1, a reduction in secretion rate is observed in DH31-stimulated MTs. Figure 1B demonstrates the use of ion-selective electrodes to measure Na+ concentrations in the secreted droplets. Treatment of DH31 on the MTs had no effect on the Na+ concentration in the secreted droplet; however, with the application of AedaeCAPA-1, the Na+ concentration in the secreted fluid was significantly increased. Additionally, compared with unstimulated controls, DH31 led to a significantly greater Na+ transport rate, whereas AedaeCAPA-1 abolished this increase in DH31-stimulated tubules (Figure 1C). Together, these sample results demonstrate how fluid secretion rates following application of diuretic and anti-diuretic hormones can be measured as well as the ion concentration within the secreted droplets. Three to six-day old females were dissected and between 23–35 MTs were analyzed for the experiment. Differences were denoted as statistically significant if p < 0.05 using a one-way ANOVA and unpaired t-tests.
The SIET was used to assess changes in Na+ transport along the rectal pad epithelia of adult female mosquitoes. Most unstimulated recta examined exhibited haemolymph-directed Na+ transport (absorption), and so only these preparations were examined. Due to variability in baseline activity, the difference in ion flux, following either saline or peptide application, relative to initial transport activity in saline was calculated. These values yield a negative change in ion flux (ion flux following treatment – ion flux in baseline conditions), although all organs remained absorptive following treatments. A leucokinin analog (Drosokinin) was used to examine changes in Na+ absorption, which resulted in a four-fold decrease in Na+ absorption compared to saline control (Figure 2).
To assess the role of a neuropeptide, pyrokinin-2 (PK2), on ileal motility, a Rhodnius prolixus analog was used, which was previously shown to activate the A. aegypti PK2 receptor, enriched in the mosquito ileum28. Relative to baseline levels, PK2 significantly inhibits ileal contractions (Figure 3).
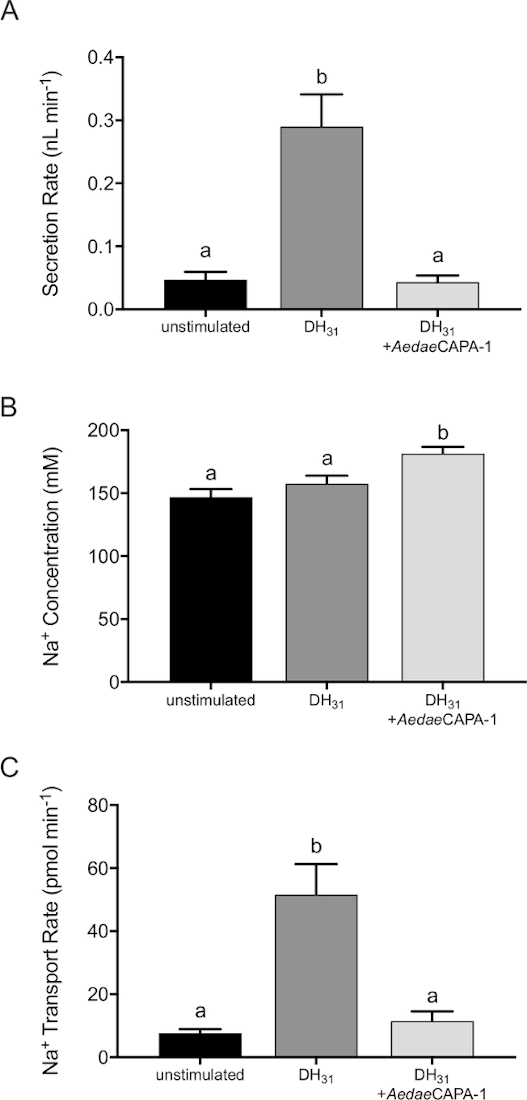
Figure 1: Effect of diuretic and anti-diuretic neuropeptides on in vitro fluid secretion rate, Na+ concentration and transport rate by adult female A. aegypti MTs. AedaeCAPA-1 (0.1 fM) was applied to MTs stimulated with DromeDH31 (25 nM). (A) MTs were incubated for 60 min with neuropeptides and 120 min for unstimulated controls. (B) Na+ concentrations in the secreted droplet were measured using ISME and values were used to calculate transport rate (C). Columns that are significantly different from controls are denoted with the same letter, as determined by a one-way ANOVA and Bonferroni post-test (n = 23–35). Please click here to view a larger version of this figure.
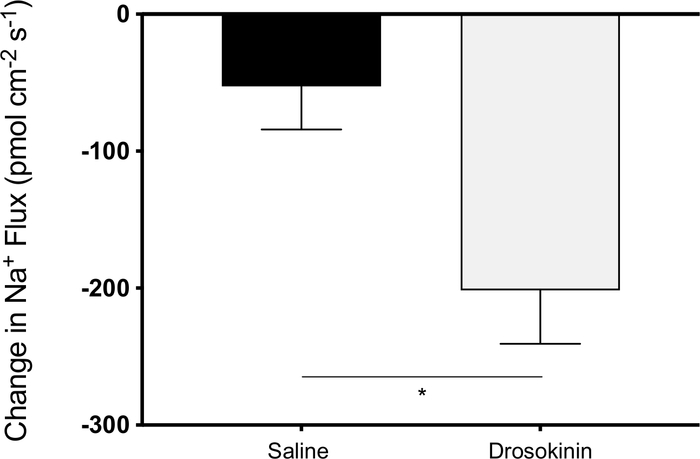
Figure 2: Changes in Na+ transport across female rectal pad epithelia measured using the SIET. Na+ absorption was recorded in unstimulated baseline saline conditions, followed by treatment with either additional saline (vehicle control) or 1 µM Drosokinin (n = 10–12). Changes in haemolymph-directed Na+ flux were measured for each treatment. There was a four-fold decrease in Na+ absorption in response to Drosokinin, as determined by an unpaired two-tailed t-test. Please click here to view a larger version of this figure.
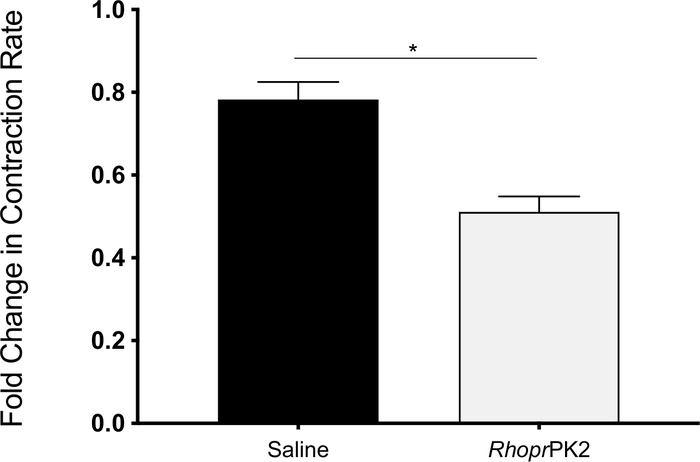
Figure 3: Change in ileal motility of female mosquitoes following saline (vehicle control) and RhoprPK2 application. The change in contraction frequency following both treatments was recorded for every sample relative to baseline conditions (n = 32). RhoprPK2 decreased ileal motility, as determined by a paired two-tailed t-test. Please click here to view a larger version of this figure.
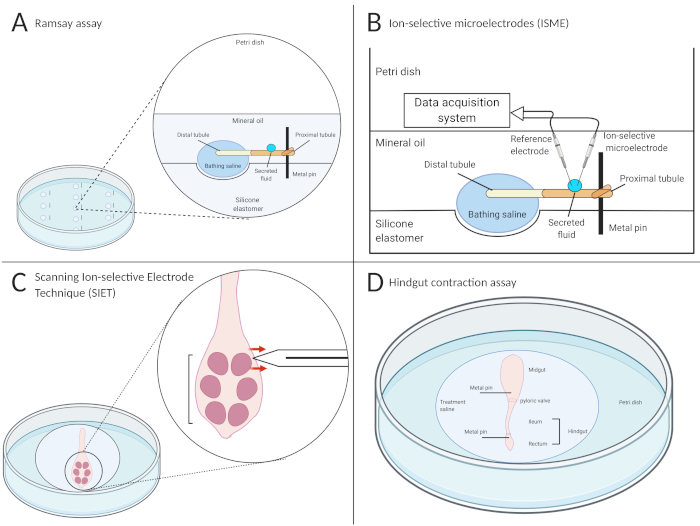
Figure 4: Schematic of the Ramsay assay, use of ion-selective microelectrodes (ISME), the scanning ion-selective electrode technique (SIET), and hindgut contraction assays. Illustrations of the four methods utilized to determine the functional roles of endocrine factors on the excretory system of the mosquito, Aedes aegypti. (A) The Ramsay assay is used to measure fluid secretion rates in isolated Malpighian tubules. The proximal (open) end of the tubule is wrapped around a pin, surrounded by mineral oil, while the distal (closed) end is bathed in the treatment saline. After a set time period, the proximal end will secrete the primary urine, accumulating as droplets on the pin. (B) A reference electrode and ISME are placed in the secreted droplet, connected to a data acquisition system, to measure ion concentration. (C) The SIET measures ion transport using an ion-selective microelectrode perpendicularly placed ~2 µm away from the organ adhered to the bottom of a poly-L-lysine dish. The direction of the red arrows in this schematic indicate that Na+ is being absorbed into the saline, indicated by a greater voltage recording along the epithelium relative to the region away from organ surface. (D) Hindgut contraction assays are conducted using video recordings to assess changes in contraction frequency, length, and relaxation duration. In this schematic, the organs are placed inside a well, and a metal pin is inserted directly into the midgut and rectum to allow the ileum to freely float and contract. Treatments are added into the well to assess changes in motility both qualitatively and quantitatively. Created using BioRender. Please click here to view a larger version of this figure.
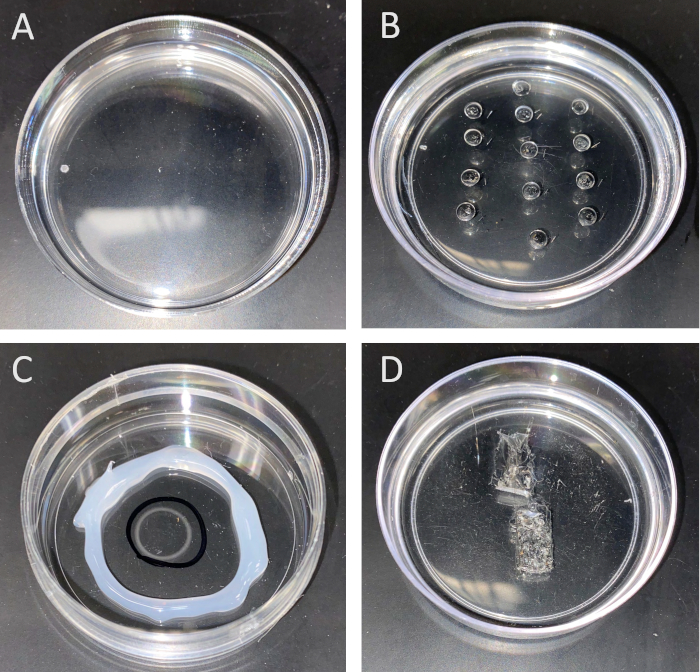
Figure 5: Dishes used for (A) dissecting mosquitoes, (B) Ramsay assay, (C) SIET and (D) contraction assay measurements. A, B and D are completely coated with silicone, whereas C is coated with poly-L-lysine in the center. To prepare the dish used for the Ramsay assay (B), a small silicone-coated Petri dish is utilized, and small ~4–5 mm wells are carved 1 cm apart (around ~20 wells can be carved/dish). Minutien metal pins are placed beside each well to hold onto the isolated tubule to measure fluid secretion rate. The poly-L-lysine dish (C) allows the organ to adhere to the bottom during SIET measurements. The wells in the contraction assay dish (D) are filled with saline and any following treatments. The contracting organ placed inside the well is observed using a video camera. Please click here to view a larger version of this figure.
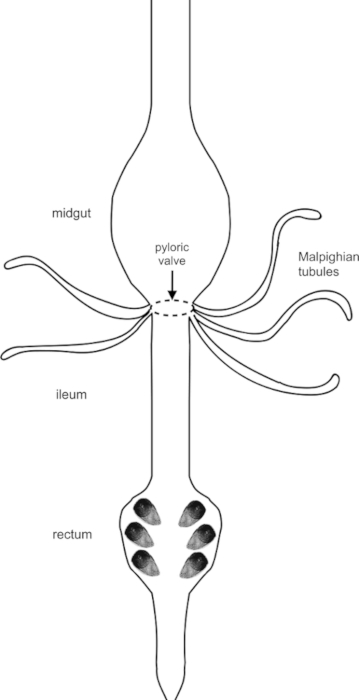
Figure 6: The excretory system of mosquito, Aedes aegypti. After a bloodmeal, solutes, water, and nutrients are passed along the alimentary canal, which includes (but is not limited to) the midgut, pyloric valve, MTs, and hindgut. The midgut is the main site for food digestion and nutrient absorption. The five MTs transport water, solutes, and waste from the surrounding haemolymph and secrete this primary urine into the hindgut, made up of an anterior ileum and posterior rectum, the latter of which includes four (male) or six (female) rectal pads. The hindgut acts as the final reabsorption site before waste is excreted. Please click here to view a larger version of this figure.
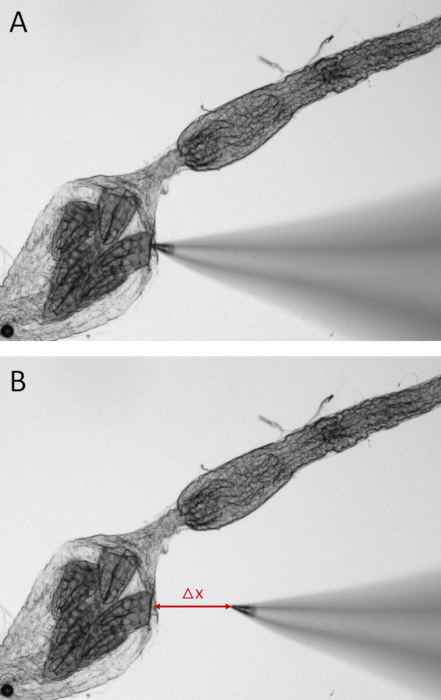
Figure 7: Still images of a dissected mosquito hindgut during SIET measurements. To measure ion activity along the epithelia of one of the six rectal pads, the Na+-selective microelectrode is placed 2 µm away from the tissue (A). Voltage is recorded at this site before the microelectrode tip is moved at an excursion distance (Δx) of 100 µm away from this point (B). The voltage gradient between these two points is used to calculate the concentration gradient and ion flux. Please click here to view a larger version of this figure.
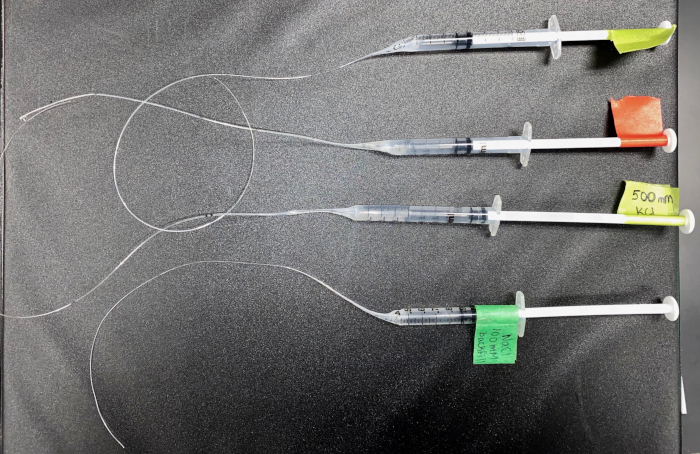
Figure 8: The backfill syringes for ISME and SIET. A picture of the syringes used to backfill NaCl and KCl solutions into the microelectrodes. This shows four 1 mL slip-tip syringes with disposable needles, with a modified stretched tip to allow for insertion into an electrode. Please click here to view a larger version of this figure.
| 2X Aedes aegypti saline | ||
| For 500 mL: | ||
| In a beaker, add 400 mL of ddH2O, then add the following solutes: | ||
| Component | Weight (g) | Final concentration (mM) |
| NaCl | 8.766 | 150 |
| HEPES | 5.957 | 25 |
| KCl | 0.253 | 3.4 |
| NaOH | 0.3 | 7.5 |
| NaHCO3 | 0.151 | 1.8 |
| MgSO4 | 0.12 | 1 |
| CaCl2-2H20 | 0.249 | 1.7 |
| Stir, and adjust pH to 7.1 | ||
| Add ddH2O to final volume of 500 mL to make 2x Aedes saline | ||
| 10X glucose | ||
| For 500 mL: | ||
| In a beaker, add 450 mL of ddH2O, then add: | ||
| Component | Weight (g) | Final concentration (mM) |
| Glucose | 4.5 | 5 |
| Stir and add ddH2O to final volume of 500 mL | ||
| To make 1X Aedes aegypti saline: | ||
| For 100 mL: | ||
| In a beaker, add 40 mL of ddH2O, then add: | ||
| Component | Volume (mL) | |
| 2x Aedes saline | 10 | |
| Stir, adjust pH to 7.1 | ||
| Filter sterilize | ||
Table 1: Making Aedes aegypti saline. To make Aedes saline, separately prepare 2X Aedes saline and 10x glucose, store in 4 °C fridge. Use these two solutions to prepare working stocks (1x Aedes saline), used for dissecting tissues, Ramsay assays, and contraction assays.
| Calcium-free Aedes aegypti Saline + NMDG | ||
| For 500 mL: | ||
| In a beaker, add 400 mL of ddH2O, then add the following solutes: | ||
| Component | Weight (g) | Final concentration (mM) |
| NaCl | 1.17 | 20 |
| NMDG | 25.38 | 130 |
| HEPES | 5.957 | 25 |
| KCl | 0.253 | 3.4 |
| NaHCO3 | 0.151 | 1.8 |
| MgSO4 | 0.12 | 1 |
| Glucose | 0.9 | 5 |
| Stir, and adjust pH to 7.1 | ||
| Add ddH2O to final volume of 500 mL | ||
Table 2: Modified Aedes aegypti saline used for SIET measurements. This Ca2+-free saline is used to prevent spontaneous hindgut contractions during SIET measurements. This saline is specific for measuring Na+ transport, as it consists of reduced Na+ (20 mM) made up by equimolar substitution with N-methyl-D-glucamine (NMDG) to reduce background noise.
| DH31-stimulated MTs | Droplet diameter (units) | Droplet diameter (um) | Droplet volume (um3) | Secretion Rate (nl/min) |
| n | Treatment #1 (60 min) | Treatment #1 (60 min) | Treatment #1 (60 min) | Treatment #1 (60 min) |
| 1 | 16 | 313.73 | 16167692.56 | 0.27 |
| 2 | 14 | 274.51 | 10831090.91 | 0.18 |
| 3 | 17 | 333.33 | 19392547.25 | 0.32 |
| 4 | 13 | 254.90 | 8671977.67 | 0.14 |
| 5 | 14 | 274.51 | 10831090.91 | 0.18 |
| 6 | 22 | 431.37 | 42029685.15 | 0.70 |
| 7 | 15 | 294.12 | 13321768.16 | 0.22 |
| 8 | 16 | 313.73 | 16167692.56 | 0.27 |
| 9 | 20 | 392.16 | 31577524.53 | 0.53 |
| Mean | 16.33333333 | 320.26 | 18776785.52 | 0.31 |
| STD ERR | 0.06 |
Table 3: Sample Ramsay assay data. Table showing sample data collected from DH31-stimulated MTs from adult female mosquitoes. When measuring the diameter of the secreted droplet, first measure with the stereomicroscope ocular micrometer and note down the value. Next, multiply the ocular unit diameter with the calibration conversion conducted before the experiment to get the diameter of the droplet in µm. Calculate the volume of the equation next, using the equation noted in step 7.7, followed by the secretion rate, using the equation noted in step 7.8.
