استكشاف ديناميكيات المادة اللينة باستخدام النيوترونات
يعد التحقيق في الخصائص الديناميكية للبروتينات والببتيدات جزءا رئيسيا من الأبحاث الفيزيائية الحيوية ، وهناك العديد من الطرق المتطورة اليوم للوصول إلى مجموعة واسعة من المناظر الطبيعية للطاقة1. إن ربط ديناميكيات البروتينات المكتشفة تجريبيا بوظيفتها البيولوجية مهمة أكثر صعوبة بكثير ، حيث تتطلب نماذج رياضية معقدة ومحاكاة ديناميكية بمساعدة الكمبيوتر. تم التأكيد على أهمية التحليل الطيفي النيوتروني لتحليل حركات البروتين في العديد من الدراسات التي لاقت استحسانا جيدا والمعترف بها على نطاق واسع1،2،3،4،5. قبل استكشاف مشهد الطاقة المتنوع لديناميكيات البروتين الداخلية ، يلزم تقديم نظرة عامة قصيرة على العمليات الديناميكية في المادة اللينة وكيف يمكن للنيوترونات الوصول إليها.
إن حساسية النيوترونات للتكوين النظيري ونوع التفاعلات التي تعرضها مع المادة اللينة تجعل تشتت النيوترون أحد أكثر تقنيات التحقيق تنوعا6. هناك مجموعة واسعة من مقاييس طول الارتباط وأوقات الارتباط التي يمكن للنيوترونات الوصول إليها ، من الإثارة النووية والاهتزازات الذرية إلى الحركات الجماعية وعمليات الاسترخاء البطيئة مثل الدوران متساوي الخواص والحركات المنتشرة. عند التحقيق في النيوترونات المتناثرة لنقل الطاقة الخاصة بها ، يمكن التمييز بين ثلاثة تفاعلات رئيسية: التشتت المرن ، حيث لا يوجد تبادل للطاقة بين النيوترون الوارد والجسيمات في العينة. التشتت غير المرن ، مع تبادل طاقة كبير قابل للقياس الكمي بين النيوترون والجسيمات ؛ والحالة الغريبة للتشتت شبه المرن الذي يعين نقل طاقة صغير جدا مقارنة بالطاقة النيوترونية الحادثة 1,7. توفر هذه التفاعلات معلومات دقيقة حول المواد التي تم التحقيق فيها وتشكل الأساس النظري لمجموعة واسعة من تقنيات تشتت النيوترونات.
في التشتت المرن ، يسجل الكاشف اتجاهات النيوترونات كنمط حيود ، والذي يوضح موقع ذرات العينة بالنسبة لبعضها البعض. يتم الحصول على معلومات حول ارتباطات المواقع الذرية (أي الكثافة المتكاملة S(Q) فيما يتعلق بنقل الزخم Q ، والذي يتعلق بالمعلومات الهيكلية وحدها). يشكل هذا المبدأ أساس الحيود النيوتروني8.
ينشأ التعقيد عندما لا يكون نقل الطاقة صفرا بسبب الإثارة والتقلبات الداخلية في مادة العينة. وهذا يشكل أساس التحليل الطيفي النيوتروني، حيث يتم التحقيق في النيوترونات المتناثرة كدالة لكل من نقل الطاقة E ونقل الزخم Q. يتم الحصول على المعلومات الديناميكية والهيكلية. يقيس التحليل الطيفي النيوتروني نفس الكثافة المتكاملة S(Q) لنقل الطاقة (أي تغير سرعة النيوترونات بسبب تشتت العينة، S(Q,ω) = S(Q, E)، والذي يشار إليه أيضا باسم عامل البنية الديناميكية)9.
لحساب التشتت من مادة ما ، من الأنسب استخدام دالة ارتباط الزوج 7,10. في حالة الحيود ، تعطي دالة ارتباط الزوج الثابت G(r) احتمال إيجاد مركز جسيم على مسافة معينة r من مركز جسيم آخر. يعمم التحليل الطيفي دالة ارتباط الزوج الثابت ويتضمن الطاقة/التردد/الوقت في معادلة التشتت. تصبح دالة ارتباط الزوج G(r) دالة للوقت G(r, t) ، والتي يمكن أن تتحلل إلى دالة ارتباط زوج ذرة متميزة G D(r ، t) ، ودالة ارتباط ذاتي GS(r, t). تصف هذه نوعين من الارتباطات: الحركات المترابطة بين الأزواج للذرات التي تحكم التشتت المتماسك ، والارتباط الذاتي الذي يحكم التشتت غير المتماسك10.
التشتت المتماسك هو التشتت من “المتوسط” ويعتمد على المرحلة النسبية للموجات المتناثرة. في نظام التشتت صغير الزاوية ، تتداخل الموجات النيوترونية المتناثرة من مراكز تشتت مختلفة (ذرات مختلفة) بشكل بناء (لها مراحل مماثلة) ، وتلاحظ الحركة الجماعية للذرات مع تعزيز كثافة قوية. يصف التشتت المتماسك بشكل أساسي تشتت نيوترون واحد من جميع النوى في العينة10.
عندما لا يحدث أي تداخل بناء بين الموجات النيوترونية المتناثرة من مراكز مختلفة ، يتم اتباع ذرة واحدة في الوقت المناسب ، ويتم ملاحظة الارتباط الذاتي بين موضع الذرة في الوقت t = 0 ونفس الذرة في الوقت t. وبالتالي ، يتم فقدان المعلومات المتعلقة بالمواقع النسبية للذرات ، وينصب التركيز فقط على التقلبات المحلية. التشتت من التقلبات المحلية يحكم التشتت غير المتماسك. التشتت غير المتماسك متساوي الخواص ، ويساهم في إشارة الخلفية ، ويحط من الإشارة إلى الضوضاء10,11.
بالجمع بين كل ما سبق ، نميز أربع عمليات تشتت نيوترونية رئيسية10: (1) متماسكة مرنة (تقيس ارتباطات المواقع الذرية) ، (2) متماسكة غير مرنة (تقيس الحركات الجماعية للذرات) ، (3) مرنة غير متماسكة (تساهم في الخلفية ، وتقلل من شدة التشتت بواسطة عامل Debye-Waller (DWF) وتقيس عامل البنية المرنة غير المتماسكة (EISF) ، واصفا هندسة الحركات المنتشرة في الهندسة المحصورة ، و (4) غير متماسك غير مرن (يقيس ديناميكيات الذرة الواحدة والارتباط الذاتي).
تتراوح العمليات الديناميكية التي يمكن للنيوترونات الوصول إليها في علم الأحياء من تخميد الاهتزازات الذرية والجزيئية منخفضة التردد ، وتفاعل جزيئات المذيبات مع الأسطح الحيوية ، وعمليات الانتشار في طبقة الترطيب للجزيئات الكبيرة والهندسة المحصورة ، إلى الحركات المنتشرة الانتقالية والدورانية والهابطة قصيرة المدى ، ومجالات البروتين والحركات الألوستريكية1 . يعتمد التنوع الواسع في طرق وأدوات النيوترون لقياس ديناميكيات البروتين على كيفية تحقيق الشعاع النيوتروني الحادث أو الصادر وكيفية إجراء تحليل الطاقة للنيوترونات المتناثرة. من المحاور الثلاثية إلى وقت الطيران والتشتت الخلفي ومطياف الصدى المغزلي ، يمكن للمرء استكشاف العمليات الديناميكية ذات الأوقات المميزة بين 1 × 10-14 ثانية و 1 × 10-6 ثانية (الفيمتو ثانية إلى ميكروثانية)12.
يحتوي مختبر أوك ريدج الوطني ، بمصدريه النيوترونيين المشهورين ، مصدر نيوترون الانتشار – SNS13 ومفاعل تدفق النظائر العالية – HFIR14 ، على واحد من أفضل أجنحة أجهزة قياس الطيف للتحقيق في الديناميكيات في المواد الحيوية. بعض الأمثلة الأكثر بلاغة تشمل استخدام مطياف المروحية النيوترونية الباردة (CNCS) في SNS15 للتحقيق في الاضطراب الديناميكي لمياه الترطيب حول بروتين الفلورسنت الأخضر في المحلول 16 أو الاهتزازات الجماعية دون بيكو ثانية للعديد من البروتينات17. تتمثل إحدى المشكلات المتكررة لتحقيقات تشتت النيوترونات غير المرنة في أن بعض العمليات البيولوجية بطيئة للغاية بحيث لا يمكن ملاحظتها. بدون إعدادات متطرفة تؤدي إلى خسارة كبيرة في كثافة النيوترون ، تقتصر مطياف وقت الطيران على دقة طاقة 10 μeV ، المقابلة لمقياس زمني أقصى يبلغ ~ 200 ps10,11. هذا لا يكفي لمراقبة الحركات واسعة النطاق في البروتينات. لذلك ، غالبا ما تكون هناك حاجة إلى أدوات ذات دقة طاقة أعلى مثل مطياف التشتت الخلفي. وقد أثبت الجمع بين تقنيات وقت الطيران والتشتت الخلفي قوته في التحقيق في التغير في الديناميكيات الداخلية للسيتوكروم P450cam (CYP101)، وهو إنزيم يحفز كافور الهيدروكسيل18.
كان الانتشار المجهري الذي تم قياسه بواسطة مطياف التشتت الخلفي في SNS-BASIS19 محددا بشكل جيد بشكل مدهش ويمكن فصله إلى انتشار الماء (الترطيب ، السيتوبلازم ، والماء الشبيه بالكتلة) وانتشار مكونات الخلايا في الديدان المسطحة المستوية ، وهو أول حي يتم دراسته بواسطة تشتت النيوترون20 . التشتت الخلفي هو تقنية طيفية عالية الدقة ، ولكنه يقتصر أيضا على عدة μeV = عدة نانو ثانية ، في حين أن الديناميكيات البطيئة في المواد الحيوية تظهر أيضا كوقت بقاء للارتباط بين الموقع الذري أو اتجاهات الدوران (على سبيل المثال ، عمليات الاسترخاء ، والتي تحدث بانتظام في النطاق الزمني من عشرة إلى مئات من النانو ثانية).
التحليل الطيفي لصدى الدوران النيوتروني (NSE) هو تقنية التشتت النيوتروني الوحيدة التي تصل إلى هذه الدقة العالية. على عكس تقنيات النيوترونات الأخرى ، لا يتطلب NSE التدرج اللوني للحزمة لأنه يستخدم المرحلة الميكانيكية الكمومية للنيوترونات ، وهي لحظاتها المغناطيسية. يسمح التلاعب باللحظات المغناطيسية باستخدام توزيع الطول الموجي العريض لحزمة النيوترون ، في حين أن التقنية حساسة لتغيرات سرعة النيوترونات الصغيرة جدا في ترتيب 1 × 10-4. تم استخدام NSE بنجاح للتحقيق في الديناميكيات البطيئة للبروتينات في محلول للعديد من البروتينات. من بين هذه الدراسات الرائدة العديدة ، نعترف بدراسة المرونة القطاعية للغلوبولين المناعي للخنزير21 ؛ حركات المجال المقترن في Taq polymerase22 ؛ حركات المجال في رباعي نازعة هيدروجيناز الكحول الخميرة23 ؛ تغيير التشكيل في كيناز الفوسفوغليسيرات عند ربط الركيزة3 ؛ تنشيط حركات المجال والانتشار الديناميكي للإشارات الألوستريك في بروتين العامل المساعد التنظيمي 1 (NHERF1)لتبادل Na + / H (NHERF1) 4،24،25 ؛ ديناميات الحالة المدمجة لاختزال أيون الزئبق26 ؛ وانتشار الهيموجلوبين في خلايا الدم الحمراء27. كشفت دراستان حديثتان في ديناميكيات البروتين عن مرونة الجسم المضاد البشري الغلوبولين المناعي G (IgG) كربيع إنتروبي28 وخصائص مساهمة المذيبات في ديناميكيات بروتين المايلين الأساسي المضطرب جوهريا (MBP)5.
تشرح هذه المقالة المبادئ الأساسية ل NSE ، والطرق التحضيرية المتعددة الموصى بها لإجراء تحقيق شامل في ديناميكيات البروتين ، بالإضافة إلى المنهجية والبروتوكول التجريبي للحصول على بيانات NSE في مطياف NSE في SNS ، SNS-NSE. يميز البروتوكول بروتينين: IgG ، وهو بروتين جسم مضاد بشري منتظم ، والبروتين المضطرب جوهريا MBP. وتناقش بإيجاز الآثار البيوفيزيائية، والأهمية البحثية للأمثلة، والقيود المفروضة على هذه التقنية.
التحليل الطيفي NSE ، طريقة قياسات الديناميكيات البطيئة
NSE هي تقنية مستقطبة تستخدم وقت الطيران النيوتروني لقياس تبادل الطاقة (فقدان الاستقطاب) بسبب التفاعل شبه المرن بين النيوترونات والذرات في العينة. في قلب التحليل الطيفي NSE يكمن مبدأان أساسيان: (1) قدرة الدوران النيوتروني على التفوق في المجال المغناطيسي بتردد يتناسب مع القوة 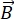 المغناطيسية ، أي تردد Larmor29 ، و (ب) صدى الدوران أو صدى هان ، الذي يمثل التلاعب وإعادة تركيز إشارة الاستقطاب عند تطبيق سلسلة من نبضات الترددات الراديوية30.
المغناطيسية ، أي تردد Larmor29 ، و (ب) صدى الدوران أو صدى هان ، الذي يمثل التلاعب وإعادة تركيز إشارة الاستقطاب عند تطبيق سلسلة من نبضات الترددات الراديوية30.
يمكن تلخيص أساسيات عملية NSE في بضع خطوات بسيطة 6,11 باستخدام الشكل 1. (1) يتم استقطاب الحزمة النيوترونية التي ينتجها المصدر (الموضع 1) (الموضع 2) ، وتوجيهها ، ونقلها (الموضع 3) ، وتصل إلى مدخل مطياف NSE ، حيث يتم تدويرها بمقدار 90 درجة بواسطة الزعانف النصف الأول من pi-half (الموضع 4). (2) يصبح الشعاع المستقطب (على سبيل المثال ، اللحظات المغناطيسية النيوترونية) عموديا على خطوط المجال المغناطيسي للمغناطيس الأول (منطقة الركود الأولى ، الموضع 5) ويبدأ في الاستباق. (3) في نهاية المغناطيس ، تتراكم الدوران النيوتروني زاوية ركود معينة تتناسب مع قوة المجال المغناطيسي ووقت الطيران الذي يقضيه في الداخل (يتناسب عكسيا بشكل أساسي مع سرعة النيوترون). يتم ترميز السرعات النيوترونية الفردية داخل زاوية الركود الخاصة بها في نهاية منطقة الركود الأولى. (4) بالقرب من موضع العينة ، يعكس pi-flipper (الموضع 6) اتجاه الدوران بمقدار 180 درجة ، مما يغير علامة زاوية الركود. (5) تتفاعل النيوترونات مع جزيئات العينة (الموضع 7) وتتناثر. (6) تدخل النيوترونات المتناثرة وتسبقها في منطقة الركود الثانية (الموضع 8) ولكنها تصبح معكوسة المنحى. (7) يستخدم زعانف نصف pi أخرى (الموضع 9) لتدوير اتجاه الدوران من الاتجاه العمودي إلى الاتجاه الأفقي. سيؤدي ذلك إلى إيقاف الركود ، وترجمة زاوية الركود φ إلى استقطاب يتناسب مع cos (φ). (8) يختار المحلل (الموضع 10) النيوترونات بناء على اتجاه واحد. إذا كان التفاعل مع العينة مرنا، فلن تتغير سرعة النيوترون. ستقضي النيوترونات قدرا مماثلا من الوقت في الطيران في منطقتي الركود الأولى والثانية ، ويتم استرداد زوايا الركود المتراكمة بالكامل. يتم استعادة الاستقطاب الكامل على الكاشف (الموضع 11) كصدى للاستقطاب الأصلي (أي صدى الدوران). (9) ومع ذلك ، في NSE ، يكون التشتت شبه مرن ، لذا فإن تبادل الطاقة الصغير بين النيوترونات وجزيئات العينة يؤدي إلى سرعات نيوترونية مختلفة بعد تشتت العينة. نظرا للسرعات المختلفة ، ستقضي النيوترونات وقتا إضافيا في الطيران عبر منطقة الركود الثانية ولن تكون قد استعادت زاوية الركود بشكل صحيح. يتم استرداد الاستقطاب الجزئي على الكاشف ، ويتناسب فقدان الاستقطاب بسبب استرخاء الدوران مع تحويل كوس-فورييه للدالة الطيفية S(Q ، ω) ، دالة التشتت الوسيطة F (Q ، t). (10) تتناسب المعلمة الزمنية للدالة F(Q, t) مع قوة المجال المغناطيسي للركود. وبالتالي ، فإن مسح فقدان الاستقطاب كدالة لقوة المجال المغناطيسي ينتج عنه وظيفة استرخاء تعتمد على العمليات الديناميكية داخل العينة.

الشكل 1: صورة فوتوغرافية لمقياس الطيف NSE في SNS (SNS-NSE) ومخطط مسار الذبابة النيوترونية مع أهم المكونات الوظيفية. من اليمين إلى اليسار: 1 = مصدر النيوترون. 2 = المروحيات-بندر-المستقطب-نظام الغالق الثانوي; 3 = أدلة النقل شعاع; 4 = pi / 2 زعانف لأول 90 درجة دوران ؛ 5 = منطقة الركود الأولى; 6 = pi flipper ل 180 ° دوران الدوران ؛ 7 = منطقة العينة وبيئة العينة (هنا ، يتم عرض فرن التبريد) ؛ 8 = منطقة الركود الثانية ؛ 9 = pi / 2 زعانف للدوران الثاني 90 درجة ؛ 10 = محلل ؛ 11 = كاشف. (لاحظ أن أجزاء من 3 ، وكذلك 2 و 1 ، تقع خلف الجدار الأزرق داخل التدريع ؛ يتم استبدال المروحيات بمحدد السرعة ل NSE القائم على المفاعل). يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
