We applied the above protocol to electroporation-mediated RNAi targeting MCO2 gene and negative control genes (EGFP for siRNA and bla for dsRNA) (i) in the abdomen of I. senegalensis (Figure 4), (ii) in the thorax of I. senegalensis (Figure 5), and (iii) in the abdomen of P. zonata (Figure 6). The results of the RNAi experiments are summarized in Table 1. Because negatively charged siRNA/dsRNA is incorporated only into positively charged cells, RNAi phenotypes were observed around the region where the positive electrode was placed for electroporation.
In both I. senegalensis and P. zonata, inhibition of melanin pigmentation (i.e., black, brown, and reddish brown) appeared in patches around the region where the positive electrode was placed (white arrowheads and dotted lines in Figure 4, Figure 5, and Figure 6) when MCO2 RNAi was performed in combination with electroporation (Table 1), as previously reported in N. pygmaea9. By contrast, no phenotypic effects were observed around the electroporation site when the control gene was injected (EGFP siRNA or bla dsRNA) (Figure 4, Figure 5, and Figure 6, Table 1). In addition, injecting the MCO2 gene without electroporation had no effect on adult pigmentation (Figure 4, Table 1), indicating that electroporation is essential for RNAi in Odonata. It should be noted that the blue, green, and yellow colorations are not affected by the RNAi of MCO2 gene that is involved in melanin synthesis in the cuticle, which plausibly reflect the fact that these body colors are attributed to pigment granules present in the epidermal cells that are visible through the transparent cuticle26. As shown in Figure 4, no remarkable phenotypic differences were recognized between the individuals subjected to siRNA treatment and dsRNA treatment, whereas considerable variation in size and location of the RNAi phenotype was observed among different individuals subjected to the same RNAi treatment (ex. compare two examples of IsMCO2 dsRNA in Figure 4).
To determine the developmental stage most suitable for the RNAi treatment, we compared the phenotypic consequences of the RNAi treatment at five morphological stages (stage A-E) in the final larval instar of I. senegalensis (Figure 7A). Inhibition of melanin pigmentation caused by MCO2 RNAi was observed in all emerged adults when injected at the stages A and B (Figure 7C). When injected at the stages C and D, suppression of melanin pigmentation was certainly observed in some emerged adults, but other adults exhibited abnormal coloration caused by wounds (Figure 7B).
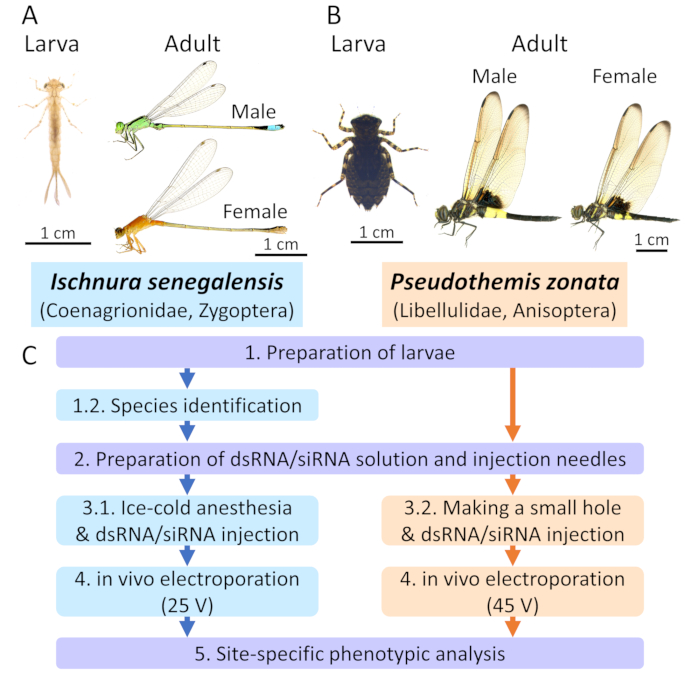
Figure 1: Electroporation-mediated RNAi methods in Odonata. A. Ischnura senegalensis (Coenagirionidae) as a representative damselfly species. B. Pseudothemis zonata (Libellulidae) as a representative dragonfly species. C. The overall scheme of the protocols. Blue and orange boxes indicate the protocols for I. senegalensis and P. zonata, respectively. The purple boxes indicate the common protocols applied to both species. Please click here to view a larger version of this figure.
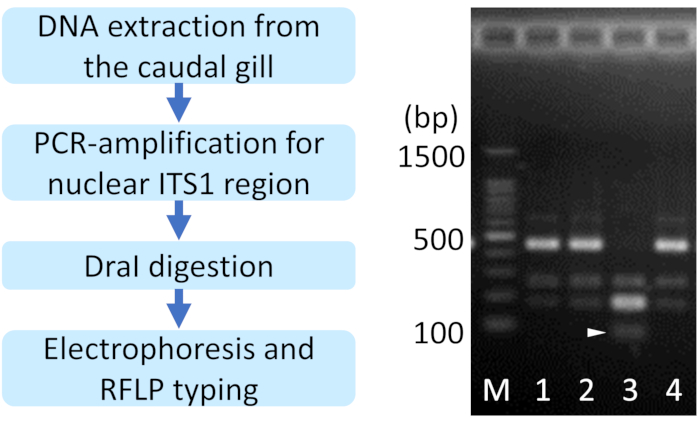
Figure 2: A representative result of restriction fragment length polymorphism (RFLP)-based species identification for Ischnura species. Arrowhead indicates the I. senegalensis-specific band. 1, 2, 4: I. asiatica, 3: I. senegalensis, M: 100-base pair ladder marker. Please click here to view a larger version of this figure.

Figure 3: Electroporation-mediated RNAi method in Odonata. A-C. Ice-cold anesthesia of I. senegalensis. Arrowheads indicate a larva. A. Putting a larva on crushed ice with a wet paper. B. Magnified view of a larva on ice. C. A larva covered with a wet paper on ice. D-G. RNAi method for I. senegalensis. D. Injection into the thorax. E. Electroporation on the thorax. F. Injection into the abdomen. G. Electroporation on the abdomen. H-J. RNAi method for P. zonata. H. Making a small hole on the abdomen. I. Injection into the abdomen. J. Electroporation on the abdomen. Arrows indicate the point of making a hole or injection. +, -: Positive/negative side electrodes. Numbers indicate the abdominal segment. Please click here to view a larger version of this figure.
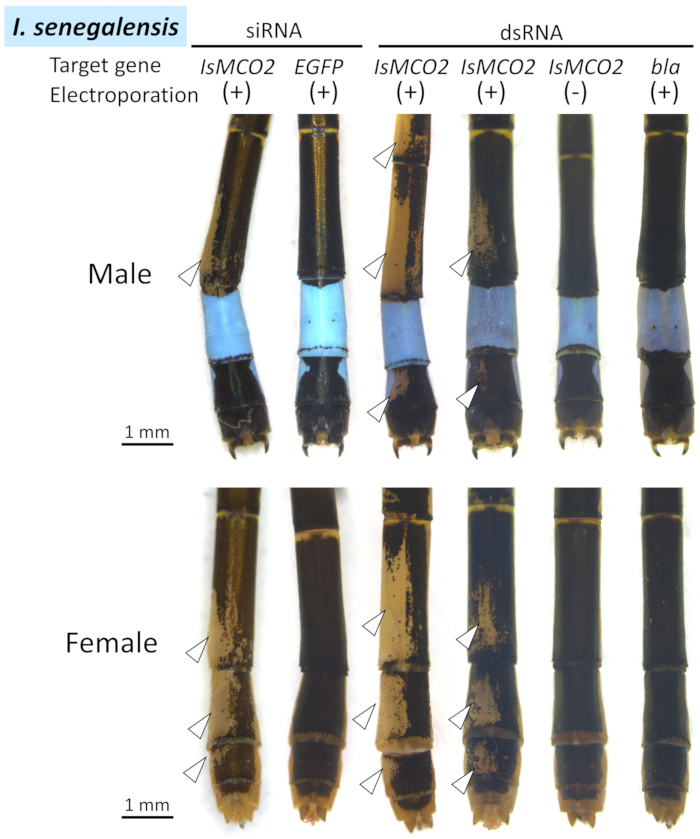
Figure 4: Dorsal views of RNAi phenotypes on the abdomen of I. senegalensis. White arrowheads indicate the regions of suppressed melanization. Please click here to view a larger version of this figure.
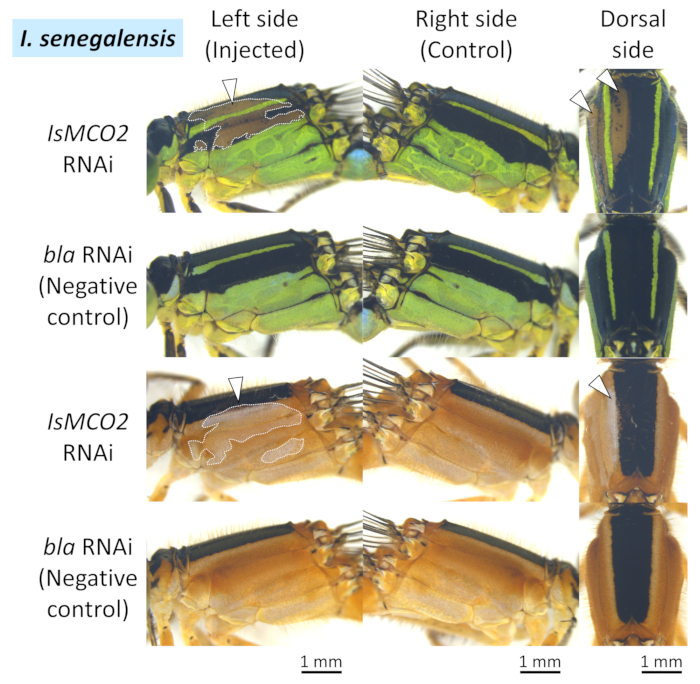
Figure 5: Lateral and dorsal views of RNAi phenotypes on the thorax of I. senegalensis. White arrowheads and dotted lines indicate the regions of suppressed pigmentation. Please click here to view a larger version of this figure.
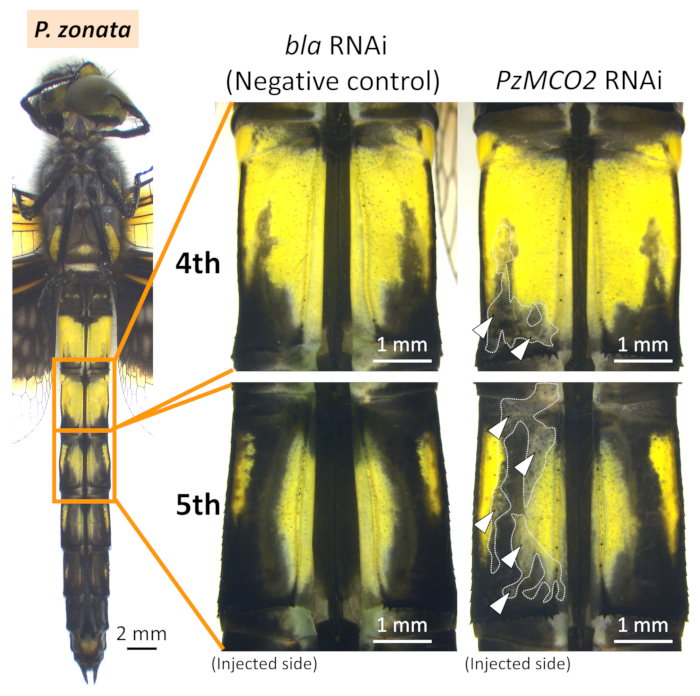
Figure 6: Ventral views of RNAi phenotypes in the abdomen of P. zonata. White arrowheads and dotted lines indicate the regions of suppressed melanization. Please click here to view a larger version of this figure.
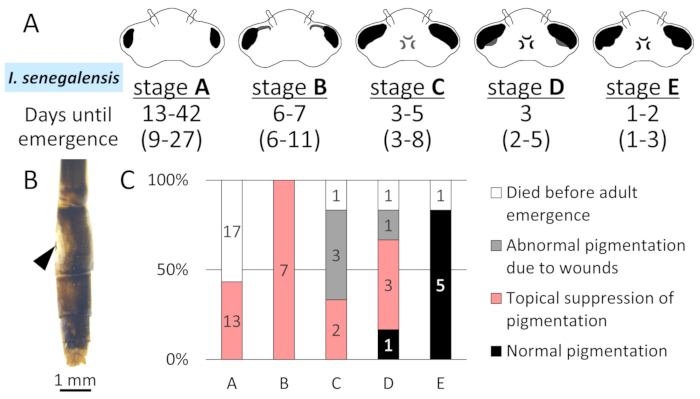
Figure 7: Stage dependent IsMCO2 RNAi effects during the final larval instar of I.senegalensis. A. Morphological changes in the compound eyes at five morphological stages (stage A-E) and the number of days to adult emergence in this study. Numbers in parentheses are from previous report14. B. Abnormal pigmentation due to wounds. Arrowhead indicates electroporation site. C. The effect of RNAi at five morphological stages on adult pigmentation in I. senegalensis. The number on the bar indicates the number of individuals. Please click here to view a larger version of this figure.
| Species | I.senegalensis | P.zonata | |||||||||
| Injected region | Abdomen | Thorax | Abdomen | ||||||||
| siRNA/dsRNA | siRNA | dsRNA | dsRNA | dsRNA | |||||||
| Target gene | IsMCO2 | EGFP | IsMCO2 | IsMCO2 | bla | IsMCO2 | bla | PzMCO2 | PzMCO2 | bla | |
| Electroporation | + | + | + | – | + | + | + | + | – | + | |
| Injected larvae | 22 | 25 | 30 | 6 | 53 | 12 | 20 | 17 | 5 | 9 | |
| Emerged adults | 7 | 6 | 13 | 4 | 40 | 11 | 14 | 11 | 2 | 5 | |
| Adults with less pigmented regions (ratio) | 7 (100%) |
0 (0%) |
13 (100%) |
0 (0%) |
0 (0%) |
10 (91%) |
0 (0%) |
11 (100%) |
0 (0%) |
0 (0%) |
|
Table 1. The effect of RNAi on adult pigmentation in I. senegalensis and P. zonata. Results at stage A are shown in I. senegalensis. IsMCO2: multicopper oxidase 2 gene of I. senegalensis, EGFP: Enhanced green fluorescent protein gene, bla: beta lactamase gene from pGEM-T Easy Vector, PzMCO2: multicopper oxidase 2 gene of P. zonata. The results for the control genes represent the total number of experiments the authors have conducted to date.
