En los últimos años, la importancia de los experimentos unicelulares se ha hecho evidente. Los datos de células individuales permiten la investigación de la variabilidad célula a célula, la resolución de correlaciones de parámetros intracelulares y la detección de cinéticas celulares que permanecen ocultas en las mediciones deconjuntos 1,2,3. Con el fin de investigar la cinética celular de miles de células individuales en paralelo, se necesitan nuevos enfoques que permitan el monitoreo de las células en condiciones estandarizadas durante un período de tiempo de varias horas hasta varios días seguido de un análisis cuantitativo de datos 4. Aquí, presentamos imágenes de células vivas de matrices unicelulares (LISCA), que combina el uso de matrices microestructuradas con microscopía de lapso de tiempo y análisis de imágenes automatizado.
Se han establecido y publicado en la literatura5,6. Aquí, describimos brevemente microescala plasma-iniciado patrón de proteínas (μPIPP). Un protocolo detallado de la fabricación de matrices unicelulares utilizando μPIPP también se encuentra en la referencia7. El uso de matrices unicelulares permite la alineación de miles de células en puntos de adhesión estandarizados que presentan microambientes definidos para cada célula y, por lo tanto, reduce una fuente de variabilidad experimental (Figura 1A). Las matrices unicelulares se utilizan para monitorear los cursos de tiempo de los marcadores fluorescentes con el propósito de indicar una variedad de procesos celulares. La microscopía a largo plazo en el modo de lapso de tiempo de barrido permite monitorear una gran área de las matrices unicelulares y, por lo tanto, tomar muestras de datos unicelulares en alto rendimiento durante un tiempo de observación de varias horas o incluso días. Esto genera pilas de línea de tiempo de imágenes de cada posición de la matriz (Figura 1B). Con el fin de reducir la gran cantidad de datos de imagen y extraer los cursos de tiempo de fluorescencia unicelular deseados de una manera eficiente, se requiere un procesamiento automatizado de imágenes que aproveche el posicionamiento de las células (Figura 1C).
El reto de LISCA es adaptar los protocolos experimentales y las herramientas computacionales para formar un ensayo de alto rendimiento que genere datos cuantitativos y reproducibles de cinética celular. En este artículo proporcionamos una descripción paso a paso de los métodos individuales y cómo se combinan en un ensayo LISCA. Como ejemplo, discutimos el curso del tiempo de la expresión mejorada de la proteína fluorescente verde (eGFP) después de la entrega artificial del mRNA. La expresión del eGFP que sigue entrega del mRNA es descrita por las ecuaciones de la tarifa de reacción que modelan la traducción y la degradación del mRNA. Ajustar la función del modelo para el curso temporal de la concentración de eGFP a la lectura LISCA de la intensidad de fluorescencia para cada célula individual a lo largo del tiempo produce las mejores estimaciones de los parámetros del modelo, como la tasa de degradación del ARNm. Como resultado representativo discutimos la eficacia de la entrega del mRNA de dos diversos agentes lípido-basados de la transfección y cómo sus distribuciones del parámetro diferencian.
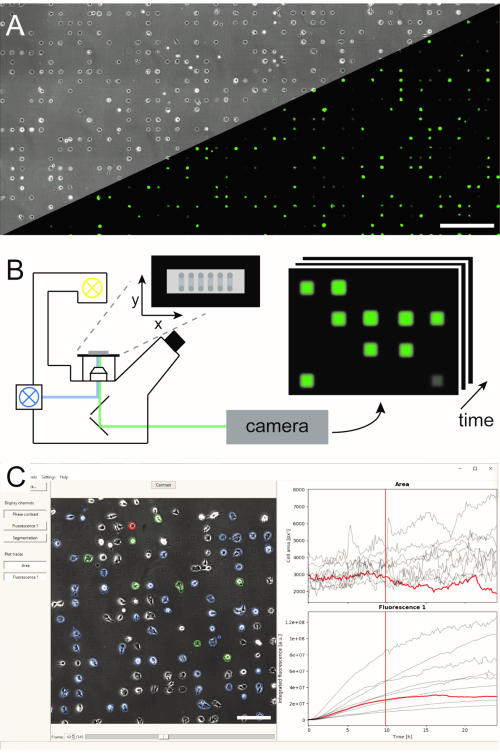
Figura 1:Representación del flujo de trabajo lisca combinando (A) micro-patrones de matrices unicelulares (B) microscopía de lapso de tiempo de barrido y (C) análisis automatizado de imágenes de series de imágenes grabadas. Las matrices unicelulares consisten en un patrón bidimensional de cuadrados adhesivos celulares con un interespacio repelente celular que conduce a una disposición de las células en el micropatrón, como se puede ver en la imagen de contraste de fase, así como la imagen de fluorescencia de las células que expresan eGFP(A). Toda el área microestructurada se toma una imagen en un modo de lapso de tiempo de escaneo tomando repetidamente imágenes en una secuencia de posiciones (B). Las series de imágenes grabadas se procesan para leer la intensidad de fluorescencia por célula a lo largo del tiempo(C). Barras de escala: 500 μm (A), 200 μm (C). Haga clic aquí para ver una versión más amplia de esta figura.
