活性氧物种(ROS)信号调节神经系统的发展和功能1。一个重要的细胞ROS来源是NADPH氧化酶(NOX),这是产生超氧化物和过氧化氢(H 2 O2)2的跨膜蛋白。NOX酶存在于整个中枢神经系统(CNS)中,NOX衍生的ROS有助于神经元发育3、4、5、6。神经干细胞的维持和分化,建立神经元极性,中性生长,突触可塑性已被证明需要足够的ROS 7,8,9,10,11水平。另一方面,NOXs不受控制地生产ROS会导致神经退行性疾病,包括阿尔茨海默氏症、多发性硬化症和创伤性脑损伤12、13、14。因此,生产生理相关的ROS对于维持健康状况至关重要。
基因编码生物传感器的开发极大地促进了细胞ROS的检测。基因编码生物传感器的一个重要优点是 ROS 信号的时间和空间分辨率增加,因为这些传感器可以专门针对不同的位置。对氧化物敏感的GFP(roGFP)是一种类型的ROS生物传感器。roGFP2-Orp1变种通过其Orp1域专门检测H2O2,这是一种来自酵母15,16的谷胱甘肽过氧化物类家庭蛋白。Orp1蛋白的氧化被转移到roGFP2,以改变其构象(图1A)。探测器在405纳米和480纳米附近显示两个兴奋峰,在515纳米处显示一个单个发射峰值。氧化后,兴奋峰周围的荧光强度会发生变化:当405 nm兴奋增加时,480纳米激发会减少。因此,roGFP2-Orp1 是一种比值生物传感器,H2O2-水平通过两个不同波长的荧光增量(图 1B)的兴奋度比率检测到。总的来说,roGFP2-Orp1 是 ROS 成像的多功能工具,可在体内高效使用。
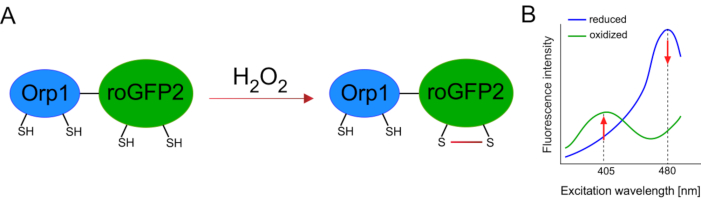
图1:roGFP2-Orp1. (A) 氧化剂转移的示意图表示和激发光谱发生在 Orp1 和 roGFP2 之间,以响应 H2O2,导致 roGFP2 的构象变化。(B) roGFP2-Orp1的激发光谱显示两个兴奋峰值在405 nm和480 nm和单个排放峰值在515 nm。H2O2氧化后,405 nm 激发增加,而 480 nm 激发减少。这会导致 H 2O 2存在的比例读出。该数字已从比兰和贝卢索夫(2017)16和摩根等人(2011)25修改。请单击此处查看此图的较大版本。
达尼奥雷里奥(斑马鱼)模型系统在应用基因编码生物传感器方面有几个优势。胚胎和幼虫的光学透明度使体内成像具有非侵入性。新的成像工具正在开发中,以实现更高的分辨率和更深的渗透17。此外,还有既定的基因操纵工具(异位mRNA表达,托尔2转基因等)和基因组编辑(塔伦斯,CRISPR/Cas9等),促进转基因动物的生成18。随着斑马鱼胚胎在母体之外发育,该系统进一步允许更容易地获取和操作胚胎。例如,单细胞阶段注射和药物治疗很容易做到。
在这里,我们使用斑马鱼通过注射体外转录的mRNA来暂时表达H2O2特异性生物传感器roGFP2-Orp1。这些胚胎可用于培养神经元的 体外 成像和 体内 成像(图2)。我们描述了从斑马鱼胚胎解剖和电镀视网膜结节细胞(RGCs)的协议,然后评估培养神经元的H2O2水平。然后,我们提出了一种利用共聚焦显微镜对罗格FP2-Orp1表达胚胎和幼虫进行 体内 成像的方法。这种方法不仅能够确定生理H2O2水平,而且还可以确定在不同的发育阶段或条件下发生的潜在变化。总的来说,该系统为检测活细胞和动物中的H2O2 提供了可靠的方法,以研究H2O2 在发育、健康和疾病中的作用。
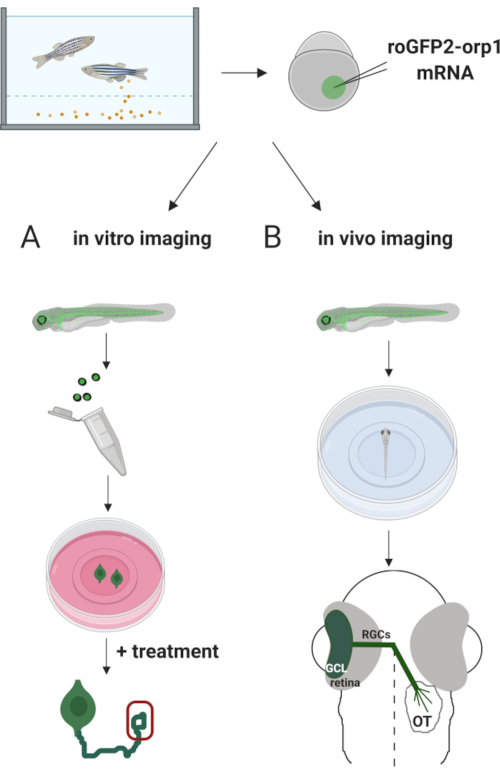
图2。实验方法大纲。简言之,胚胎采集后,roGFP2-Orp1 mRNA被注射到单细胞阶段斑马鱼胚胎的蛋黄中。发育中的胚胎可用于体外和体内成像的(A)和B。(A) GFP阳性胚胎用于解剖视网膜,用于在34马力下采集RGC。分离的 RGC 镀在 ZFCM (+) 介质中的 PDL/层膜盖上。生长锥成像可以进行RGC延长他们的轴突后,6-24小时的电镀。细胞可以接受不同的治疗,以测量H2O2水平的潜在变化。在这里,我们测量了RGC(红色)生长锥中的H 2 O2水平。(B) GFP阳性胚胎用于体内成像。在理想年龄,胚胎可以麻醉并安装在35毫米玻璃底盘上,用于共焦成像。在这里,胚胎被安装在腹膜成像的通风装置上。示意图显示斑马鱼的视网膜发育。RGC 形成结膜细胞层 (GCL),这是网状网膜的最内层。RGC轴突发展成视神经,穿过中线,形成视基亚西姆。然后,RGC轴突生长,在中脑的光学构造中形成突触。请单击此处查看此图的较大版本。
