A sinalização reativa da espécie oxigênio (ROS) regula o desenvolvimento e o funcionamento do sistema nervoso1. Uma importante fonte ros celular são as oxidases NADPH (NOX), que são proteínas transmembranas que geram superóxido e peróxido de hidrogênio (H2O2)2. Enzimas NOX são encontradas em todo o sistema nervoso central (CNS), e ros derivado do NOX contribuem para o desenvolvimento neuronal3,4,5,6. A manutenção e diferenciação das células-tronco neurais, o estabelecimento da polaridade neuronal, do crescimento do neurite e da plasticidade sináptica têm sido mostrados como necessários níveis adequados de ROS7,8,9,10,11. Por outro lado, a produção descontrolada de ROS por NOXes contribui para distúrbios neurodegenerativos, incluindo doença de Alzheimer, esclerose múltipla e lesão cerebral traumática12,13,14. Por isso, a produção de ROS fisiologicamente relevante é fundamental para a manutenção de condições saudáveis.
O desenvolvimento de biosensores geneticamente codificados facilitou muito a detecção de ROS celular. Uma vantagem importante dos biosensores geneticamente codificados é o aumento da resolução temporal e espacial do sinal ROS, pois esses sensores podem ser especificamente direcionados a locais distintos. O GFP sensível ao redox (roGFP) é um tipo desses biosensores ROS. A variante roGFP2-Orp1 detecta especificamente H2O2 através de seu domínio Orp1, que é uma proteína geneatoxitina de glutationa peroxiredoxina a partir da levedura15,16. A oxidação da proteína Orp1 é transferida para roGFP2 para alterar sua conformação (Figura 1A). A sonda exibe dois picos de excitação perto de 405 nm e 480 nm, e um único pico de emissão de 515 nm. Após a oxidação, a intensidade da fluorescência em torno dos picos de excitação muda: enquanto a excitação de 405 nm aumenta, a excitação de 480 nm diminui. Assim, roGFP2-Orp1 é um biosensor ratiométrico, e osníveisde H2O 2 são detectados pela razão de intensidades de fluorescência animadas em dois comprimentos de onda diferentes(Figura 1B). No geral, roGFP2-Orp1 é uma ferramenta versátil para imagens ROS que pode ser utilizada de forma eficiente in vivo.
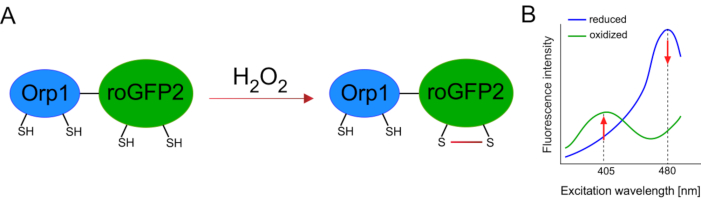
Figura 1: Espectros de representação e excitação esquemática de roGFP2-Orp1. (A) A transferência oxidante ocorre entre Orp1 e roGFP2 em resposta a H2O2, levando a alterações conformais no roGFP2. (B) O espectro de excitação do roGFP2-Orp1 exibe dois picos de excitação de 405 nm e 480 nm e pico de emissão única de 515 nm. Após a oxidação por H2O2,a excitação de 405 nm aumenta enquanto a excitação de 480 nm diminui. Isso resulta em uma leitura ratiométrica para a presença de H2O2. O número foi modificado de Bilan e Belousov (2017)16 e Morgan et al. (2011)25. Clique aqui para ver uma versão maior desta figura.
O sistema modelo Danio rerio (zebrafish) tem várias vantagens para aplicar biosensores geneticamente codificados. A transparência óptica dos embriões e larvas permite imagens in vivo não invasivas. Novas ferramentas de imagem estão sendo desenvolvidas para alcançar maior resolução e penetração mais profunda17. Além disso, existem ferramentas estabelecidas para manipulação genética (expressão ectópica mRNA, transgênese Tol2, etc.) e edição de genomas (TALENs, CRISPR/Cas9, etc.), que promove a geração de animais transgênicos18. À medida que os embriões de zebrafish se desenvolvem fora da mãe, este sistema permite ainda mais acesso e manipulação dos embriões. Por exemplo, injeções de estágio de uma célula e tratamentos medicamentosos podem ser facilmente feitos.
Aqui, usamos zebrafish para expressar transitoriamente o biosensor H2O2-específico roGFP2-Orp1 injetando mRNA in vitro transcrito. Esses embriões podem ser usados tanto para imagens in vitro de neurônios cultivados quanto para imagens in vivo (Figura 2). Descrevemos um protocolo para dissecar e emplacar células de gânglios de retina (RGCs) de embriões de zebrafish seguidos pela avaliação dos níveis de H2O2em neurônios cultivados. Em seguida, apresentamos um método para imagens in vivo de embriões e larvas que expressam roGFP2-Orp1 usando microscopia confocal. Esta abordagem não só permite determinar os níveis fisiológicos H2O2,mas também possíveis mudanças ocorridas em diferentes estágios ou condições de desenvolvimento. No geral, este sistema fornece um método confiável para detectar H2O2 em células vivas e animais para estudar o papel de H2O2 no desenvolvimento, saúde e doenças.
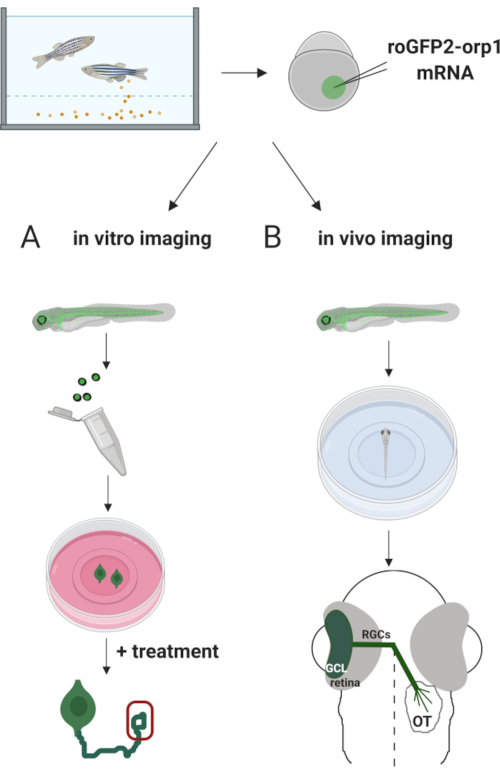
Figura 2. Esboço da abordagem experimental. Brevemente, após a coleta de embriões, roGFP2-Orp1 mRNA é injetado na gema de embriões de zebrafish estágio de uma célula. Embriões em desenvolvimento podem ser usados paraimagens in vitro ede vivo . ( A )Osembriões GFP positivos são usados para dissecar retinas para coleta de RGC a 34 hpf. Os RGCs dissociados são emplacados em tampas revestidas de PDL/laminina na mídia ZFCM (+). A imagem do cone de crescimento pode ser conduzida à medida que os RGCs estendem seus axônios após 6-24 h de revestimento. As células podem ser submetidas a diferentes tratamentos para medir as possíveis mudanças nos níveis de H2O2. Aqui, medimos h2O2 -níveisnos cones de crescimento de RGCs (vermelho). (B) Os embriões positivos GFP são utilizados para imagens in vivo. Na idade desejada, os embriões podem ser anestesiados e montados em pratos de fundo de vidro de 35 mm para imagens confocal. Aqui, os embriões são montados ventrally para imagem da retina. Esquema mostra desenvolvimento de retina em zebrafish. Os RGCs formam camada de células de glion (GCL), que é a camada mais interna da retina. Os axônios RGC desenvolvem-se em nervo óptico para cruzar a linha média, formando quiasmo óptico. Em seguida, os axônios RGC crescem dorsais para fazer sinapses no tectum óptico no cérebro médio. Clique aqui para ver uma versão maior desta figura.
