Передача сигналов активных форм кислорода (АФК) регулирует развитие и функционирование нервнойсистемы1. Важным клеточным источником АФК являются NADPH оксидазы (NOX), которые представляют собой трансмембранные белки, образующие супероксид и перекись водорода (H2O2)2. Ферменты NOX обнаруживаются по всей центральной нервной системе (ЦНС), а АФК, полученная изNOX,способствует развитию нейронов3,4,5,6. Было показано, что поддержание и дифференцировка нервных стволовых клеток, установление полярности нейронов, роста нейритов и синаптической пластичности требуют адекватных уровней АФК7,8,9,10,11. С другой стороны, неконтролируемое производство АФК ТОКС способствуют нейродегенеративным расстройствам, включая болезнь Альцгеймера, рассеянный склероз и черепно-мозговую травму12,13,14. Следовательно, производство физиологически значимого АФК имеет решающее значение для поддержания здоровых условий.
Разработка генетически закодированных биосенсоров значительно облегчила обнаружение клеточного АФК. Одним из важных преимуществ генетически закодированных биосенсоров является повышенное временное и пространственное разрешение сигнала АФК, поскольку эти датчики могут быть специально нацелены на различные места. Окислительно-чувствительный GFP (roGFP) является одним из типов таких биосенсоров ROS. Вариант roGFP2-Orp1 специфически обнаруживает H2O2 через свой домен Orp1, который представляет собой белок семейства глутатиона пероксиредоксинов из дрожжей15,16. Окисление белка Orp1 переносится в roGFP2 для изменения его конформации(рисунок 1A). Зонд демонстрирует два пика возбуждения около 405 нм и 480 нм и один пик излучения при 515 нм. При окислении интенсивность флуоресценции вокруг пиков возбуждения изменяется: при увеличении возбуждения 405 нм снижается возбуждение на 480 нм. Таким образом, roGFP2-Orp1 является логометрическим биосенсором, а уровниH2O2-определяются соотношением интенсивностей флуоресценции, возбуждаемых на двух разных длинах волн(рисунок 1B). В целом, roGFP2-Orp1 является универсальным инструментом для визуализации АФК, который можно эффективно использовать in vivo.
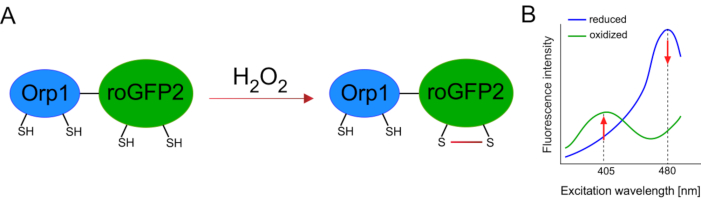
Рисунок 1:Схематическое представление и спектры возбуждения roGFP2-Orp1. (A)Перенос окислителя происходит между Orp1 и roGFP2 в ответ наH2O2,что приводит к конформационным изменениям roGFP2. (B)Спектры возбуждения roGFP2-Orp1 демонстрируют два пика возбуждения при 405 нм и 480 нм и одинарный пик излучения при 515 нм. При окислении наH2O2 возбуждение405 нм увеличивается, а возбуждение 480 нм уменьшается. Это приводит к ратиометрическому считываю для присутствия H2O2. Рисунок был изменен с Bilan and Belousov (2017)16 и Morgan et al. (2011)25. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Модельная система Danio rerio (рыбки данио) имеет ряд преимуществ для применения генетически закодированных биосенсоров. Оптическая прозрачность эмбрионов и личинок позволяет проводить неинвазивную визуализацию in vivo. Разрабатываются новые инструменты визуализации для достижения более высокого разрешения и более глубокого проникновения17. Кроме того, существуют установленные инструменты для генетических манипуляций (эктопическая экспрессия мРНК, трансгенез Tol2 и др.) и редактирования генома (TALENs, CRISPR/Cas9 и др.), что способствует генерации трансгенных животных18. Поскольку эмбрионы рыбок данио развиваются вне матери, эта система дополнительно облегчает доступ и манипулирование эмбрионами. Например, инъекции на одноклеточной стадии и медикаментозное лечение могут быть легко сделаны.
Здесь мы использовали рыбок данио для временной экспрессииH2O2-специфическогобиосенсора roGFP2-Orp1 путем инъекции транскрибированной in vitro мРНК. Эти эмбрионы могут быть использованы как для визуализации культивированных нейронов in vitro, так и для визуализации in vivo (рисунок 2). Мы описываем протокол для рассечения и покрытия ганглиозных клеток сетчатки (RGC) из эмбрионов рыбок данио с последующей оценкой уровнейH2O2в культивированных нейронах. Затем мы представляем метод визуализации in vivo roGFP2-Orp1-экспрессирующих эмбрионов и личинок с использованием конфокальной микроскопии. Такой подход позволяет не только определить физиологические уровниH2O2,но и потенциальные изменения, происходящие на разных стадиях развития или в различных условиях. В целом, эта система обеспечивает надежный метод обнаруженияH2O2 в живых клетках и животных для изучения ролиH2O2 в развитии, здоровье и болезни.
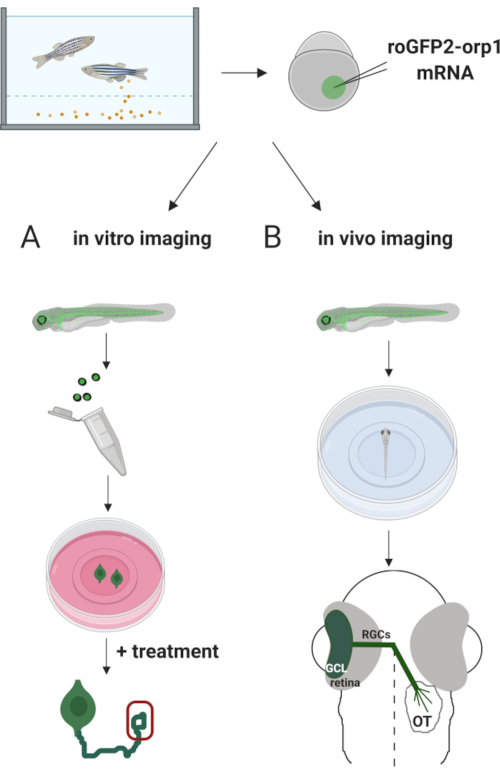
Рисунок 2. Схема экспериментального подхода. Вкратце, после сбора эмбрионов, мРНК roGFP2-Orp1 вводят в желток эмбрионов одноклеточной стадии данио. Развивающиеся эмбрионы могут быть использованы как для(A) in vitro, так и(B) in vivo визуализации. (A) GFP-положительные эмбрионы используются для рассечения сетчатки для сбора RGC при 34 л.с. Диссоциированные RGC покрываются на обшивки с PDL/ламининым покрытием в средах ZFCM (+). Визуализация конуса роста может проводиться по мере того, как RGC расширяют свои аксоны после 6-24 ч покрытия. Клетки могут подвергаться различным методам лечения для измерения потенциальных изменений в уровняхH2O2. Здесь мы измерилиH2O2-уровни в ростовых конусах RGC (красный). (B)GFP-положительные эмбрионы используются для визуализации in vivo. В желаемом возрасте эмбрионы могут быть обезболиваться и устанавливаться на стеклянную посуду со стеклянным дном 35 мм для конфокальной визуализации. Здесь эмбрионы устанавливаются вентрально для визуализации сетчатки. Схема показывает развитие сетчатки у рыбок данио. RGC образуют ганглионный клеточный слой (GCL), который является самым внутренним слоем в сетчатке. Аксоны RGC развиваются в зрительный нерв, пересекая среднюю линию, образуя зрительный хиазм. Затем аксоны RGC растут дорсально, чтобы сделать синапсы в зрительном тектуме в среднем мозге. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
