Glutamate uptake is one of the most important mechanisms controlling neurotransmission in the brain. The hippocampus, specifically, is a critical place in glutamate signaling, being an important hub connecting memory, cognition, and emotions in the brain. Following the protocol, adult male Wistar rats were used to generate representative results. Animals were anesthetized using isoflurane 3% until unconscious. After dissecting the brain, hippocampi were removed and placed in the chopper table perpendicularly to the blade. Three hundred µm thick slices were obtained at a rate of 60 per minute and collected to a petri dish containing ice-cold saline solution. Slices were separated each in a well of a 24-well plate and L-[3H]-Glutamate uptake was carried out for 5 min (unless stated otherwise) in sodium-free or regular HBSS, at 4 or 37 °C, respectively.
After an overnight period in lysis solution at 4 °C, samples were mixed with scintillation liquid and read in a scintillation counter for 60 s. The results are given in DPM and the value of the uptake carried out in sodium-free HBSS is deducted from the value obtained with regular HBSS. One DPM corresponds to 0.000000000450 or 4,5 x 10-10 Curie (Ci). By knowing the specific radioactivity of the L-[3H]-Glutamate, is possible to establish a relationship between the result in DPM and the amount of L-[3H]-Glutamate captured by each sample.
First, we carry out the uptake assay using 200 and 300 μm slices to demonstrate that the total protein amount does not influence the uptake parameter. No differences in glutamate uptake in 5 min were observed comparing slices with 200 and 300 μm (data not shown). Accordingly, the researcher has to use slices obtained from the same hippocampal region of all animals to prevent size bias. In summary, this protocol highlights the usual thickness (300 µm) used in different experiments with slices from hippocampus, striatum or cortex10,34,35. Moreover, in order to clarify if there was any difference in L-[3H]-Glutamate uptake in the three main hippocampal regions, dorsal, intermediate and ventral regions were assayed separately. There was no difference in L-[3H]-Glutamate between the three regions (Figure 3A, p = 0.638, one-way ANOVA of repeated measures). This is not surprising since the experiments were performed with naïve animals. However, the results may vary between regions when considering different experimental models or pharmacological challenges.
To make sure that the experiment was not carried out in a saturation range, the uptake was performed for 2.5, 5, 10 or 15 min. The results demonstrate that in a 10 min incubation time the amount of L-[3H]-Glutamate taken up was significantly higher when compared with 5 min (Figure 3B, p = 0.0061, F = 6.864, one-way ANOVA of repeated measures). Conversely, 10 and 15 min uptake results were not statistically different, suggesting a plateau was reached. This data shows that a 5 min incubation is adequate to measure the uptake in most conditions.
To demonstrate that the method is sensitive enough to respond to a blockage in glutamate uptake, 100 µM dihydrokainic acid (DHK), a specific GLT-1 blocker, was administered to the AHS, 5 min prior the uptake. DHK was supplied in stabilization media during 5 min previous to media replacement and [3H]L-Glutamate uptake start36, according to the following timeline: t= 0: slice stabilization in 37 °C HBSS, t= 10 min: addition of DHK to specific slices, t= 15 min: substitution of stabilization media by fresh 37 °C HBSS and beginning of [3H]L-Glutamate uptake. As expected, DHK was able to decrease glutamate uptake (Figure 3C, p = 0.0034, r squared = 0.9069, paired Student's t test) in the protocol conditions, indicating that other pharmacological or genetic manipulations can be used to modulate glutamate uptake.
As a way to determine if the samples are stable enough to withstand a delay in the measurement, some replicates were kept with no scintillation liquid at 4 °C (Figure 3D) or frozen for 7 days (Figure 3E), having the scintillation of their counterparts measured in the next day of the experiment. The data shows that there was no difference between the DPM values of the samples tested 24 h after the experiment and the samples kept refrigerated or frozen for 7 days (p = 0.229 and p = 0.9623, respectively, paired Student's t test).
D-[3H]-Aspartate is also a substrate to glutamate transporters and represents an index of glutamate uptake, since it is metabolized in a lesser extent than L-[3H]-Glutamate. The accumulation of D-[3H]-Aspartate in the slice would provide another important index of transporters activity. Therefore, the uptake of D-[3H]-Aspartate in 300 μm hippocampal slices was estimated for 2.5 and 5 min in the same conditions than L-[3H]-Glutamate. Different time points for the uptake of L-[3H]-Glutamate and D-[3H]-Aspartate under basal conditions did not present significant differences comparing both groups (Figure 3D; p = 0.0632 for radiolabeled substrate, 2-way ANOVA followed by Tukey).
Finally, to demonstrate that unlabeled glutamate in incubation media could hamper the radiolabeled transport activity the uptake buffer was supplemented with different concentrations of unlabeled glutamate (0, 50, 100 and 200 µM). Here, in Figure 3G it is demonstrated that the presence of unlabeled glutamate in incubation media significantly impact the L-[3H]-Glutamate uptake in a concentration-dependent manner in 300 μm hippocampal slices (p = 0.02, repeated measures ANOVA).
| Reagent | Final concentration (mM) |
| CaCl2 | 1.26 |
| D-Glucose | 5.55 |
| KCl | 5.36 |
| KH2PO4 | 0.44 |
| MgCl2 | 0.49 |
| MgSO4 | 0.41 |
| NaCl | 137 |
| NaHCO3 | 4.17 |
| Na2HPO4 | 0.63 |
Table 1. Hank's Balanced Salt Solution composition
| Reagent | Final concentration (mM) |
| CaCl2 | 1.26 |
| D-Glucose | 5.55 |
| Glucamine-HCl | 137 |
| pH 7.4 | |
| (adjust with concentrated HCl) | |
| Glucamine-HEPES | 4.17 |
| pH 7.4 | |
| (adjust pH with HEPES free acid 2 M) | |
| KCl | 5.36 |
| KH2PO4 | 0.44 |
| MgCl2 | 0.49 |
| MgSO4 | 0.41 |
Table 2. Sodium-Free HBSS composition
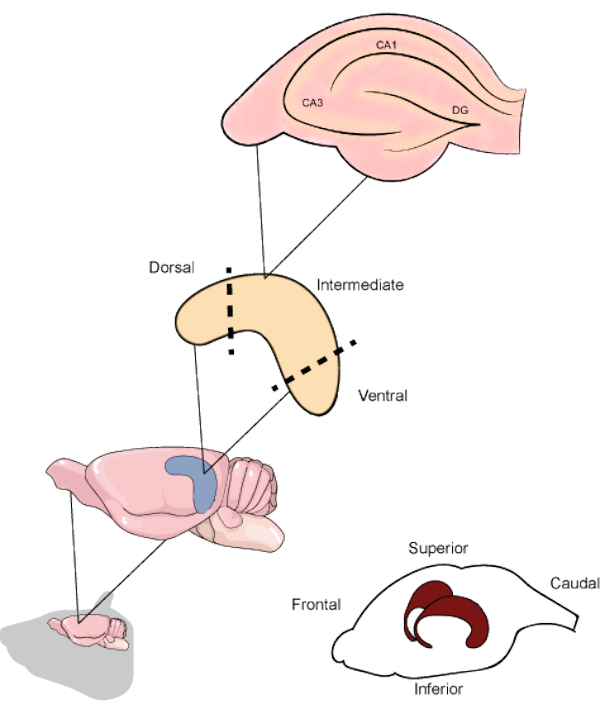
Figure 1. Schematic representation of the rodent brain and hippocampi location beneath the temporoparietal and occipital cortices. When resected, the hippocampus can be divided into the dorsal, intermediate, and ventral parts, the first being oriented in the upper direction of the brain. The hippocampal slices along the transverse axis contain the main regions (CA1, CA3 and DG). CA1: Cornu ammonis 1, CA3: Cornu ammonis 3 and DG: dentate gyrus. Please click here to view a larger version of this figure.
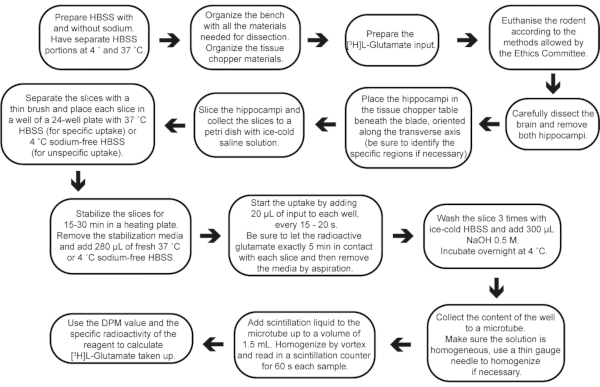
Figure 2: Step-by-step flow of the experiment, from buffer preparation to result calculations. Please click here to view a larger version of this figure.
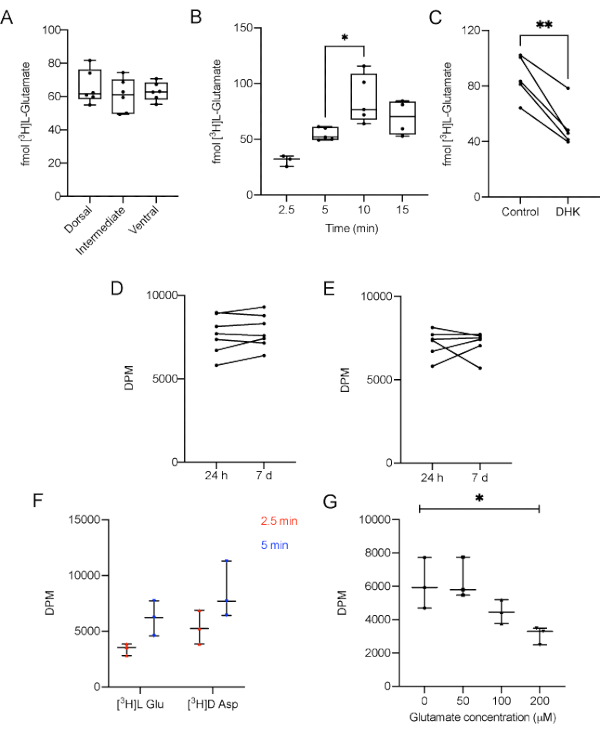
Figure 3: Representative results of L-[3H]-Glutamate uptake performed in AHS of adult male Wistar rats. A) Different regions take up the same amount of L-[3H]-Glutamate under basal conditions (p = 0.638, one-way ANOVA of repeated measures). B) The contact of L-[3H]-Glutamate with the slices for 10 min increases the amount of L-[3H]-Glutamate captured, compared with 5 min (p = 0.02, F = 6.864, one-way ANOVA). C) Dihydrokainic acid (DHK), a specific GLT-1 blocker, decreases glutamate uptake (p = 0.0034, r squared = 0.9069, paired Student's t test). Sample stability was measured after 7 days. Replicates were kept D) at 4 °C (p = 0.229, paired Student's t test) and E) -20 °C (p = 0.9623, paired Student's t test) and compared to its counterparts measured 24 h after the experiment. F) Different time points for the uptake of L-[3H]-Glutamate and D-[3H]-Aspartate under basal conditions (p = 0.0632 for radiolabeled substrate, 2-way ANOVA followed by Tukey); G) Different concentrations of unlabeled glutamate (0, 50, 100 and 200 µM) supplemented in uptake buffer (p = 0/02, one-way ANOVA of repeated measures). GLT-1: Glutamate transporter type 1. DPM: disintegrations per minute. The data in A and B are expressed as minimum value, median and maximum value. The data in C, D, and E represent paired slices, therefore they are unique values.Please click here to view a larger version of this figure.