Las bacterias utilizan varias moléculas de señalización pequeñas para adaptarse a entornos en constante cambio1,2. Por ejemplo, los autoinductores, las lactonas de N-acilhomoserina y sus oligopéptidos modificados, median la comunicación intercelular entre las bacterias para coordinar el comportamiento de la población, un fenómeno conocido como detección de quórum2. Otro grupo de moléculas de señalización pequeñas son los NSM, incluidos el ampliamente estudiado monofosfato de adenosina cíclico (cAMP), el di-AMP cíclico, el monofosfato de di-guanosina cíclico (di-GMP cíclico) y los pentafosfatos de guanosina y tetra (p)ppGpp1. Las bacterias producen estos NSM como respuesta a una variedad de diferentes condiciones de estrés. Una vez producidas, estas moléculas se unen a sus proteínas diana y regulan varias vías fisiológicas y metabólicas diferentes para hacer frente a las tensiones encontradas y mejorar la supervivencia bacteriana. Por lo tanto, la identificación de las proteínas diana es un requisito previo inevitable para descifrar las funciones moleculares de estas pequeñas moléculas.
La última década ha sido testigo de un auge del conocimiento de estas pequeñas moléculas de señalización, principalmente debido a varias innovaciones técnicas que dieron a conocer las proteínas objetivo de estas pequeñas moléculas. Estos incluyen la técnica del compuesto de captura3,4,5y la acción capilar radial diferencial del ensayo de ligando (DRaCALA)6 que se discutirá en este artículo.
Inventado por Vincent Lee y sus compañeros de trabajo en 20116,DRaCALA despliega la capacidad de una membrana de nitrocelulosa para secuestrar diferencialmente ligandos libres y unidos a proteínas. Las moléculas como las proteínas no pueden difundirse en una membrana de nitrocelulosa, mientras que los ligandos pequeños, como los NSM, pueden hacerlo. Al mezclar el NSM(por ejemplo,ppGpp) con la proteína a probar y detectarlos en la membrana, se pueden esperar dos escenarios (Figura 1): Si (p)ppGpp se une a la proteína, el radiomarcado (p)ppGpp será retenido en el centro del punto por la proteína y no se difundirá hacia afuera, dando un punto pequeño intenso (es decir, señal radiactiva fuerte) bajo un fosforrimager. Sin embargo, si (p)ppGpp no se une a la proteína, se difundirá libremente hacia afuera para producir una gran mancha con una señal radiactiva de fondo uniforme.
Además, DRaCALA puede detectar la interacción entre una molécula pequeña y una proteína no purificada en un lisato de células enteras si la proteína está presente en una cantidad suficiente. Esta simplicidad permite el uso de DRaCALA en la identificación rápida de objetivos de proteínas mediante el uso de una biblioteca de expresiones ORFeome. De hecho, las proteínas diana de cAMP7, cíclico di-AMP8, cíclico di-GMP9,10y (p)ppGpp11,12,13 se han identificado sistemáticamente mediante el uso de DRaCALA. Este artículo en vídeo utiliza (p)ppGpp como ejemplo para demostrar y describir los pasos y consideraciones críticos para realizar una proyección exitosa de DRaCALA. Cabe destacar que se recomienda leer una descripción más completa de DRaCALA14 en combinación con este artículo antes de realizar DRaCALA.
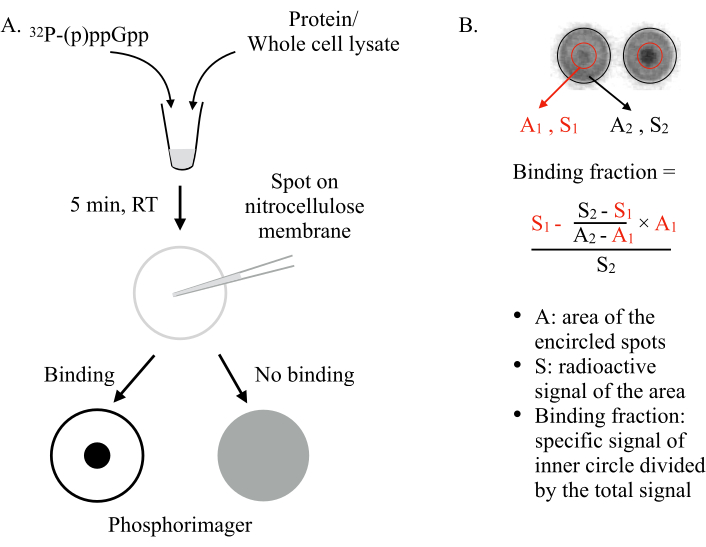
Figura 1: El principio de DRaCALA. (A) Esquema del ensayo DRaCALA. Consulte el texto para obtener más información. (B) Cuantificación y cálculo de la fracción de enlace. Consulte el texto para obtener más información. Brevemente, las manchas DRaCALA se analizarán dibujando dos círculos que circunscriben toda la mancha y el punto oscuro interno(es decir,el (p) ppGpp retenido debido a la unión de la proteína probada). La señal de unión específica es la señal radiactiva del círculo interior (S1) después de restar la señal de fondo no específica (calculada por A1 × ((S2-S1)/(A2-A1))). La fracción de unión es la señal de unión específica dividida por la señal radiactiva total (S2). Abreviaturas: DRaCALA = Acción Capilar Radial Diferencial del Ensayo de Ligandos; p)ppGpp = pentafosfatos de guanosina y tetrafosfatos; RT = temperatura ambiente. Haga clic aquí para ver una versión más grande de esta figura.
El trabajo está respaldado por una subvención del proyecto NNF (NNF19OC0058331) a YEZ, y el programa de investigación e innovación Horizonte 2020 de la Unión Europea en virtud del acuerdo de subvención Marie Skłodowska-Curie (Nº 801199) a MLS.

