Uno dei principali ostacoli nello studio delle malattie fibrotiche è la mancanza di modelli di tessuto 3D umano rappresentativi che forniscano informazioni sul comportamento dei fibroblasti e dei loro derivati patologici. Per studiare i processi fibrotici, i sistemi di coltura 2D standard sono sub-ottimali poiché i fibroblasti isolati si transdifferenziano rapidamente in miofibroblasti che esprimono α-liscia di actina muscolare (SMA) quando coltivati su substrati 2D non conformi1,2,3. Pertanto, i fibroblasti nella coltura 2D standard non riflettono un fenotipo tissutale “sano” regolare3,4,5,6. Sono state introdotte colture su substrati flessibili per simulare ambienti tissutali non fibrotici (10 kPa) e fibrotici (35 kPa)7, ma questi mancano della terza dimensione, che è molto importante per quanto riguarda la fisiopatologia. L’ingegneria tissutale offre l’opportunità di superare questa limitazione consentendo la coltura di fibroblasti in un contesto di matrice extracellulare (ECM) definito e sperimentalmente sintonizzabile, ad esempio mediante alterazioni della cellularità, della composizione ECM e della concentrazione di ECM, che possono determinare la biomeccanica tissutale.
Vari modelli 3D sono stati generati utilizzando fibroblasti. I dischi galleggianti e le microsfere sono stati tra i primi e dimostrano che il collagene viene rimodellato e compattato in modo dipendente dal tempo. I fibroblasti esercitano forze di trazione sulle fibrille di collagene, un processo che può essere facilitato dall’aggiunta di agenti pro-fibrotici come il fattore di crescita trasformante-beta 1 (TGF-β1)8,9,10,11,12,13,14,15,16. Tuttavia, le colture liberamente fluttuanti non consentono il carico esterno controllato e, pertanto, costituiscono modelli in continua contrazione o compattazione. I tessuti ingegnerizzati simili a fogli hanno aperto la possibilità di studiare la regolazione omeostatica delle proprietà biomeccaniche dei tessuti, in particolare attraverso test di deformazione uni, bi, multiassiale o ciclica17,18,19,20. Questi modelli sono stati utilizzati, ad esempio, per dimostrare l’influenza del numero di cellule sulla rigidità del tessuto, che è risultata correlata positivamente con l’integrità del citoscheletro e la contrattilità del citoscheletro dell’actomiosina.18,19. Tuttavia, è importante notare che le conversioni forza-deformazione sono complicate dalla deformazione tissutale non uniforme attorno ai punti di serraggio dei trasduttori di forza e dei punti di ancoraggio. Questa limitazione intrinseca può essere aggirata da ossa di cane o tessuti a forma di anello, offrendo una certa applicazione dei tessuti nei punti di ancoraggio21,22,23. I tessuti a forma di anello possono essere preparati distribuendo un idrogel di collagene cellulare in stampi a forma di anello. Mentre l’idrogel si compatta, un tessuto si forma attorno all’asta interna incomprimibile dello stampo, che offre resistenza per un’ulteriore contrazione del tessuto24,25,26,27. Dopo la compattazione iniziale e tipicamente massima, i tessuti possono anche essere trasferiti a distanziali regolabili per trattenere ulteriormente l’ECT circolare a una lunghezza di tessuto definita.3,24,25,26,27,28,29,30. Le proprietà biofisiche possono essere valutate in dispositivi standard orizzontali o verticali di deformazione-sollecitazione con celle di carico appropriate sotto deformazione unidirezionale o dinamica3. Poiché i tessuti hanno una struttura circolare ampiamente uniforme e possono essere tenuti su barre/ganci (punti di ancoraggio e/o trasduttori di forza), sebbene questi possano ancora racchiudere aree di compressione attorno alle barre di carico, questo formato consente una variazione di deformazione più uniforme rispetto al serraggio3. Inoltre, i tessuti ancorati suscitano una forma cellulare bipolare e le cellule si adattano alle forze tissutali per allungamento lungo le linee di forza promuovendo la trazione anisotropica.31,32,33,34,35,36. In precedenza abbiamo applicato ECT a forma di anello da fibroblasti cardiaci (CF) di ratto e umani attorno a un singolo polo rigido in esperimenti di stress funzionale e abbiamo eseguito studi di guadagno e perdita di funzione utilizzando fibroblasti trasdotti viralmente24,25,26 e studi farmacologici37. Inoltre, potremmo identificare le differenze sessuali nella fibrosi cf-mediata nel modello ECT27.
Il seguente protocollo per la generazione di ECT umano, esemplificato con CF umano primario ottenuto come CF crioconservato da fornitori commerciali (vedi Tabella dei materiali), combina i vantaggi dei tessuti a forma di anello con un modo semplice e veloce di produrre tessuti macroscopici per una piattaforma a 48 pozzi progettata per test paralleli ad alto contenuto.
È importante sottolineare che il modello ECT non è limitato a un tipo specifico di fibroblasti, con l’uso documentato nello studio di altri fibroblasti, ad esempio fibroblasti cutanei38,39. Inoltre, i fibroblasti delle biopsie del paziente funzionano altrettanto bene e la scelta dei fibroblasti dipende in ultima analisi dalla questione scientifica da affrontare.
La piattaforma utilizzata per la generazione di ECT descritta in questo protocollo è una piastra di coltura cellulare/tissutale 3D a 48 pozzetti disponibile in commercio (Figura 1A). Vengono descritti i metodi per la preparazione, la coltivazione e il monitoraggio della formazione e della funzione dell’ECT sotto una geometria definita e un carico meccanico con l’aiuto della piastra a 48 pozzetti. Gli ECT formati sono tenuti da pali flessibili integrati e il carico meccanico può essere messo a punto in base allo scopo finale utilizzando pali con diversa durezza (shore A valore 36-89), influenzandone le rigidità di flessione. Si raccomandano pali con una riva Un valore di 46. Il protocollo è, inoltre, compatibile con uno stampo circolare personalizzato precedentemente descritto, in cui l’ECT è tenuto attorno a una singola asta rigida37. Le dimensioni di questo stampo sono indicate nella Figura 1B.
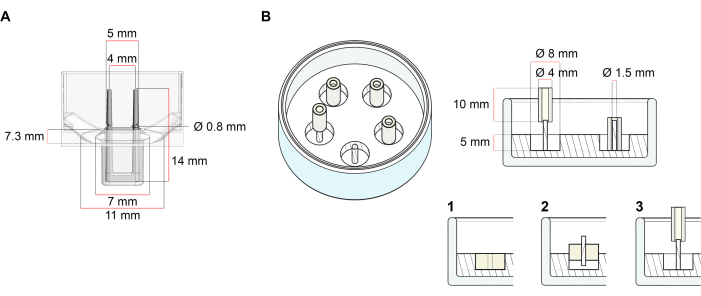
Figura 1: Rappresentazione schematica degli stampi di fusione. (A) Disegno tecnico e dimensioni di uno stampo di colata con due poli flessibili. Lo stampo comprende una circonferenza interna delimitata da una parete corta che contiene doppi pali di ritenzione sul corpo principale dello stampo. I poli flessibili hanno una distanza orizzontale libera l’uno dall’altro e sono collegati alla base. Lo stampo consente un volume di fusione di 180 μL. Il pozzetto di ogni stampo consente una capacità volumetrica di almeno 600 μL di terreno di coltura. Diverse composizioni di materiali possono essere utilizzate per produrre pali con rigidità specifiche (ad esempio, TM5MED-TM9MED). (B) Disegno tecnico e dimensioni di uno stampo a forma di anello con una singola asta rigida. Si tratta di uno stampo alternativo con geometria e ambiente meccanico distinti, che può essere utilizzato con il protocollo di fusione ECT37. Il metodo di assemblaggio dello stampo a forma di anello è stato adattato dai formati più grandi pubblicati28,41. In breve, il metodo include (1) l’imprinting di distanziali di stampaggio in politetrafluoroetilene (PTFE) (diametro 8 mm) in polidimetilsilossano (PDMS, silicone) versato in piatti di vetro (diametro 60 mm) e (2) il fissaggio di un supporto per palo PDMS (diametro 1,5 mm) concentricamente all’interno della cavità cava formata, che serve a (3) tenere un palo rimovibile (tubo in silicone di 4 mm di diametro). Lo spazio vuoto risultante consente 180 μL di volume di colata. Ogni piatto di vetro può contenere più stampi impressi (mostrati in modo esemplare con 5 stampi) e ha la capacità di raggiungere fino a 5 ml di terreno di coltura. Fare clic qui per visualizzare una versione più grande di questa figura.
