Die Modellierung der amyotrophen Lateralsklerose (ALS) im Labor ist aufgrund der überwiegend sporadischen Natur von über 80 % der Fälle1 in Verbindung mit der großen Anzahl genetischer Mutationen, von denen bekannt ist, dass sie krankheitserregend sind2, eine einzigartige Herausforderung2. Trotzdem haben alle Fälle von ALS das verbindende Merkmal gemeinsam, dass es vor der vollständigen neuronalen Degeneration zu einer dysfunktionalen Kommunikation zwischen präsynaptischen Motoneuronen und postsynaptischen Muskelzellen kommt3,4. Klinisch, wenn Patienten die Konnektivität der verbleibenden oberen und unteren Motoneuronen verlieren, weisen sie während der gesamten Krankheit Merkmale der neuronalen Hyper- und Hypoerregbarkeit auf5,6,7,8,9, die komplexe zugrunde liegende molekulare Veränderungen dieser Synapsen widerspiegeln, die wir als ALS-Forscher zu verstehen versuchen.
Mehrere transgene Modelle haben gezeigt, dass Verschlechterung und Desorganisation der neuromuskulären Verbindung mit der Expression von ALS-verursachenden genetischen Mutationen auftreten, einschließlich SOD110, FUS11,12, C9orf7213,14,15,16 und TDP4317,18,19 durch morphologische Bewertungen, einschließlich der Bewertung von synaptischen Boutons, Wirbelsäulendichten und prä- / postsynaptischer Organisation. Mechanistisch war es seit den bahnbrechenden Arbeiten von Cole, Hodgkin und Huxley in den 1930er Jahren auch möglich, synaptische Reaktionen durch elektrophysiologische Techniken entweder in vitro-Zellkultur oder in Gewebeschnittpräparaten zu bewerten20. Durch diese Strategien haben viele ALS-Modelle synaptische Übertragungsdefizite gezeigt. Zum Beispiel verursacht eine mutierte Variante von TDP43 eine erhöhte Feuerfrequenz und verringert die Aktionspotentialschwelle in NSC-34 (Rückenmark x Neuroblastom Hybridzelllinie 34) motorisch-neuronenähnlichen Zellen21. Dieselbe Variante verursacht auch eine dysfunktionale synaptische Übertragung an der neuromuskulären Verbindung (NMJ) vor dem Auftreten von verhaltensbedingten motorischen Defiziten in einem Mausmodell22. Es wurde zuvor gezeigt, dass die mutierte FUS-Expression in einem Drosophila-Modell von FUS-ALS vor lokomotorischen Defekten zu einer reduzierten synaptischen Übertragung am NMJ führt11. Ein kürzlich veröffentlichter Bericht unter Verwendung induzierter pluripotenter Stammzellen, die aus C9orf72-Expansionsträgern gewonnen wurden, zeigte eine Verringerung des leicht freisetzbaren Pools synaptischer Vesikel23. Insgesamt unterstreichen diese und andere Studien, wie wichtig es ist, ein umfassenderes Verständnis der Mechanismen aufzubauen, die der synaptischen Signalübertragung in krankheitsrelevanten Modellen der ALS zugrunde liegen. Dies wird entscheidend sein, um die Pathobiologie von ALS zu verstehen und potenzielle therapeutische Ziele für Patienten zu entwickeln.
Methoden der Strom- und Spannungsspannzellen waren von unschätzbarem Wert bei der Bestimmung von Membraneigenschaften wie Leitwert, Ruhemembranpotential und Quantengehalt einzelner Synapsen20,24. Eine der wesentlichen Einschränkungen der Elektrophysiologie besteht jedoch darin, dass sie technisch anspruchsvoll ist und nur Erkenntnisse von einem einzelnen Neuron gleichzeitig liefert. Die konfokale Lebendzellmikroskopie, gekoppelt mit spezifischen fluoreszierenden Sonden, bietet die Möglichkeit, die synaptische Übertragung von Neuronen räumlich-zeitlich zu untersuchen25,26,27. Obwohl dieser Fluoreszenzansatz kein direktes Maß für die neuronale Erregbarkeit ist, kann er eine relative Messung von zwei molekularen Korrelationen der synaptischen Funktion liefern: synaptische Vesikelfreisetzung und Calciumtransienten an synaptischen Terminals.
Wenn ein Aktionspotential die präsynaptische terminale Region von Neuronen erreicht, werden Calciumtransienten ausgelöst, was den Übergang von einem elektrischen Signal zum Prozess der Neurotransmitterfreisetzung erleichtert28. Spannungsgesteuerte Kalziumkanäle, die in diesen Bereichen lokalisiert sind, regulieren die Kalziumsignalisierung streng, um die Kinetik der Neurotransmitterfreisetzung zu modulieren29. Die ersten berichteten fluoreszenzbasierten Aufzeichnungen von Calciumtransienten wurden entweder mit dem Zweiwellenlängenindikator Fura-2 AM oder dem Einzelwellenlängenfarbstoff Fluo-3 AM30,31,32 durchgeführt. Während diese Farbstoffe zu dieser Zeit großartige neue Erkenntnisse boten, leiden sie unter mehreren Einschränkungen wie unspezifischer Kompartimentierung innerhalb von Zellen, aktivem oder passivem Farbstoffverlust aus markierten Zellen, Photobleichen und Toxizität, wenn sie über längere Zeiträume abgebildet werden33. In den letzten zehn Jahren sind genetisch kodierte Kalziumindikatoren zu den Arbeitspferden für die Abbildung verschiedener Formen neuronaler Aktivität geworden. Diese Indikatoren kombinieren ein modifiziertes fluoreszierendes Protein mit einem Calciumchelatorprotein, das nach der Bindung von Ca2+-Ionen schnell die Fluoreszenzintensität wechselt34. Die Anwendung dieser neuen Indikatoren ist enorm und ermöglicht eine viel einfachere Visualisierung intrazellulärer Calciumtransienten sowohl in vitro- als auch de vivo-Umgebungen. Eine Familie dieser genetisch kodierten Reporter, bekannt als GCaMP, wird jetzt breit genutzt. Diese Indikatoren enthalten eine C-terminale Calmodulindomäne, gefolgt von grün fluoreszierendem Protein (GFP), und sind durch eine N-terminale Calmodulin-Bindungsregion begrenzt35,36. Die Calciumbindung an die Calmodulindomäne löst eine Wechselwirkung mit der Calmodulin-bindenden Region aus, was zu einer Konformationsänderung der gesamten Proteinstruktur und einer erheblichen Erhöhung der Fluoreszenz der GFP-Einheit führt35,36. Im Laufe der Jahre hat diese Reporterfamilie mehrere Entwicklungen durchlaufen, um unterschiedliche Auslesungen für bestimmte Kalziumtransienten mit spezifischer Kinetik (langsam, mittel und schnell) mit jeweils leicht unterschiedlichen Eigenschaften zu ermöglichen37,38. Hier wurde die Verwendung des Reporters GcaMP6 hervorgehoben, von dem zuvor gezeigt wurde, dass er einzelne Aktionspotentiale und dendritische Calciumtransienten in Neuronen sowohl in vivo als auch in vitro nachweist37.
Calciumtransienten in der präsynaptischen Region lösen synaptische Vesikelfusionsereignisse aus, die eine Freisetzung von Neurotransmittern in die Synapse und die Initiierung von Signalereignissen in der postsynaptischen Zelle verursachen28,39. Synaptische Vesikel werden sowohl schnell freigesetzt als auch recycelt, da die Zelle homöostatisch eine stabile Zellmembranoberfläche und einen leicht lösbaren Pool fusionsfähiger membrangebundener Vesikel beibehält40. Der hier verwendete Styrylfarbstoff hat eine Affinität zu Lipidmembranen und verändert seine Emissionseigenschaften spezifisch anhand der Ordnung der umgebenden Lipidumgebung41,42. Somit ist es ein ideales Werkzeug für die Markierung von synaptischen Recyclingvesikeln und die anschließende Verfolgung dieser Vesikel, da sie später nach neuronaler Stimulation freigesetzt werden41,42. Das Protokoll, das generiert und optimiert wurde, ist eine Adaption der ursprünglich von Gaffield und Kollegen beschriebenen Konzepte, die es uns ermöglicht, styrylfarbstoffmarkierte synaptische Vesikelpunkta im Laufe der Zeit kontinuierlich zu visualisieren41.
Hier werden zwei verwandte fluoreszenzbasierte Methoden beschrieben, die zuverlässig über spezifische zelluläre Ereignisse berichten, die an der synaptischen Übertragung beteiligt sind. Protokolle wurden definiert, um die Dynamik des depolarisationsvermittelten präsynaptischen terminalen Kalziumeinflusses und der synaptischen Vesikelexozytose in kultivierten Neuronen zu untersuchen. Hier konzentrieren sich Methoden und repräsentative Ergebnisse auf die Verwendung primärer kortikaler oder motorischer Neuronen von Nagetieren als In-vitro-Modellsystem, da es veröffentlichte Studien mit diesen Zelltypen gibt43,44. Diese Methoden sind jedoch auch auf differenzierte humane i3-kortikal-ähnliche Neuronen anwendbar45, da wir auch mit beiden Protokollen in derzeit laufenden Experimenten in unserem Labor erfolgreich waren. Das allgemeine Protokoll ist in einem schrittweisen linearen Format dargestellt (siehe Abbildung 1). Kurz gesagt, um die Kalziumdynamik in Neuriten zu untersuchen, werden reife Neuronen mit Plasmid-DNA transfiziert, um den fluoreszierenden Reporter GCaMP6m unter einem Cytomegalovirus (CMV) -Promotor zu exprimieren37,46. Transfizierte Zellen haben eine geringe basale grüne Fluoreszenz, die in Gegenwart von Kalzium zunimmt. Interessante Regionen werden angegeben, um Fluoreszenzänderungen während unserer Manipulation zu überwachen. Damit lassen sich stark räumlich und zeitlich begrenzte Schwankungen des Calciums messen37,46. Zur Beurteilung der synaptischen Vesikelfusion und -freisetzung werden reife Neuronen mit Styrylfarbstoff beladen, der in synaptische Vesikelmembranen eingebaut wird, während sie in präsynaptischen Zellen recycelt, reformiert und mit Neurotransmittern wieder geladen werden41,42,43,47,48. Die aktuellen Farbstoffe, die zu diesem Zweck verwendet werden, markieren synaptische Vesikel entlang von Neuriten und werden als Proxy für diese Regionen in Live-Imaging-Experimenten verwendet, wie die gemeinsame Färbung von Styrylfarbstoff und Synaptotagmin durch Kraszewski und Kollegen gezeigt wurde49. Hier sind repräsentative Bilder ähnlicher Färbungen enthalten, die ebenfalls durchgeführt wurden (Abbildung 2A). Frühere Forscher haben solche Farbstoffe ausgiebig verwendet, um die Dynamik synaptischer Vesikel an der neuromuskulären Verbindung und den Hippocampus-Neuronen zu melden48,49,50,51,52,53,54,55,56 . Durch die Auswahl punktueller Regionen von farbstoffbeladenen Vesikeln und durch die Überwachung der Abnahme der Fluoreszenzintensität nach der Vesikelfreisetzung können die funktionelle synaptische Übertragungskapazität und die zeitliche Dynamik der Freisetzung nach Stimulation untersucht werden43. Für beide Methoden wird ein Medium verwendet, das eine hohe Konzentration an Kaliumchlorid enthält, um Zellen zu depolarisieren, um neuronale Aktivität nachzuahmen. Bildgebungsparameter werden angegeben, um Intervalle von unter einer Sekunde zu erfassen, die sich über eine Basisnormalisierung erstrecken, gefolgt von unserer Stimulationserfassungsperiode. Fluoreszenzmessungen zu jedem Zeitpunkt werden bestimmt, auf den Hintergrund normalisiert und über den experimentellen Zeitraum quantifiziert. Calcium-Influx-vermittelte GCaMP6m-Fluoreszenzerhöhung oder effektive synaptische Vesikel-Exozytose-Styryl-Farbstofffreisetzungsfluoreszenzabnahme kann durch diese Strategie nachgewiesen werden. Detaillierter methodischer Aufbau und Parameter für diese beiden Protokolle und eine Diskussion über ihre Vorteile und Grenzen werden im Folgenden beschrieben.
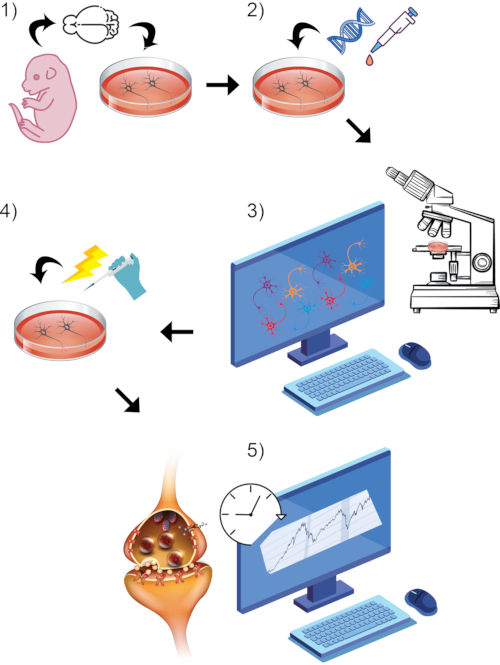
Abbildung 1: Visuelles Rendering des allgemeinen Protokollprozesses. (1) Isolieren und kultivieren Sie primäre Nagetierneuronen in vitro zum gewählten Reifungszeitpunkt. (2) Einführung von GCaMP-DNA oder Styrylfarbstoff als Reporter der synaptischen Aktivität. (3) Richten Sie das Bildgebungsparadigma mit einem mit Live-Imaging ausgestatteten Konfokalmikroskop und der zugehörigen Software ein. Beginnen Sie mit dem Baseline-Aufzeichnungszeitraum. (4) Während Zellen noch live aufgenommen werden, stimulieren sie Neuronen durch hohe KCl-Badperfusion. (5) Bewertung von Fluoreszenzintensitätsmessungen im Laufe der Zeit, um Calciumtransienten oder synaptische Vesikelfusion zu messen. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
