Неспецифический эндоцитарный путь макропиноцитоза позволяет клеткам усваивать различные внеклеточные компоненты, включая питательные вещества, белки, антигены и патогены, путем массового поглощения внеклеточной жидкости и ее составляющих1. Хотя это важно для биологии многочисленных типов клеток, все чаще описывается, что путь макропиноцитоза играет важную роль в биологии опухоли, где благодаря макропиноцитарному поглощению опухолевые клетки способны выживать и размножаться в присутствии истощенного питательными веществами микросреды2,3. Поглощение внеклеточных макромолекул, включая альбумин и внеклеточный матрикс, а также некротический клеточный мусор, обеспечивает альтернативный источник питательных веществ для производства биомассы путем создания аминокислот, сахаров, липидов и нуклеотидов через макропиносомы и лизосомы, опосредованные слиянием, катаболизм груза4,5,6,7,8.
Индукция и регуляция макропиноцитоза сложны и могут варьироваться в зависимости от клеточного контекста. До сих пор было идентифицировано несколько индукторов макропиноцитоза, которые включают лиганды, такие как эпидермальный фактор роста (EGF), тромбоцитарный фактор роста (PDGF), галектин-3 и Wnt3A9,10,11,12,13. Кроме того, условия культивирования, которые имитируют микроокружение опухоли, могут вызвать активацию пути. Опухоли аденокарциномы протоков поджелудочной железы (PDAC) лишены питательных веществ, особенно для аминокислоты глутамина, которая заставляет как раковые клетки, так и связанные с раком фибробласты (CAF) полагаться на макропиноцитоз для выживания7,13,14,15. Кроме того, опухолевые стрессы, такие как гипоксия и окислительный стресс, могут активировать этот путь очистки16. В дополнение к многочисленным внешним инфлюенсорам, которые могут индуцировать макропиноцитоз, различные внутриклеточные пути контролируют образование макропиносом. Онкогенная Ras-опосредованная трансформация достаточна для инициирования макропиноцитарного механизма, и несколько типов рака проявляют онкогенный Ras-управляемый конститутивный макропиноцитоз4,5,9,17. В качестве альтернативы, была идентифицирована активация Ras дикого типа и Ras-независимые пути для активации макропиноцитоза в раковых клетках и CAFs10,11,15,18. Использование различных моделей in vitro в сочетании с ингибиторами лечения привело к идентификации нескольких модуляторов макропиноцитоза, которые включают натриево-водородные обменники, малый GTPase Rac1, фосфоинозитид-3-киназу (PI3K), p21-активированную киназу (Pak) и AMP-активированную протеинкиназу (AMPK)4,13,15 . Однако, учитывая множество описанных факторов и условий, регулирующих макропиноцитоз, вполне возможно, что многие другие модуляторы и стимулы остаются неоткрытыми. Идентификация новых модуляторов и стимулов может быть облегчена автоматизированной оценкой множества условий в одном эксперименте. Эта методология может пролить свет на факторы, участвующие в образовании макропиносом, и может позволить обнаружить новые малые молекулы или биологические препараты, которые нацелены на этот путь.
Здесь мы адаптировали наш ранее установленный протокол для определения степени макропиноцитоза в раковых клетках in vitro к формату микропластин с 96 лунками и автоматической визуализации и количественной оценке19,20. Этот протокол основан на флуоресцентной микроскопии, которая стала стандартом в этой области для определения макропиноцитоза in vitro и in vivo4,5,6,7,9,10,11,12,13,15,16,17,18, 19,20,21,22. Макропиносомы можно отличить от других эндоцитарных путей благодаря их способности усваивать крупные макромолекулы, такие как высокомолекулярный декстран (т.е. 70 кДа)2,3,4,20,21,22,23. Таким образом, макропиносомы могут быть определены путем поглощения внеклеточного вводимого флуорофором меченого декстрана 70 кДа. В результате макропиноцитарные везикулы проявляются в виде внутриклеточных скоплений флуоресцентных пункт с размерами от 0,2-5 мкм. Эти пункты могут быть микроскопически изображены и впоследствии количественно определены для определения степени макропиноцитоза в клетке – «макропиноцитарного индекса».
В этом протоколе описаны основные этапы визуализации макропиносом в адгезивных клетках in vitro на 96-луночной микропластине и крышках с использованием стандартного лабораторного оборудования (рисунок 1). Кроме того, приведены указания по автоматизации получения изображения и количественной оценки макропиноцитарного индекса с помощью многомодового пластинчатого считывателя клеточной визуализации. Эта автоматизация сокращает время, затраты и усилия по сравнению с нашими ранее описанными протоколами19,20. Кроме того, он позволяет избежать непреднамеренно предвзятого получения и анализа изображений и тем самым повышает воспроизводимость и надежность. Этот метод может быть легко адаптирован к различным типам клеток или считывателям пластин или использоваться для определения альтернативных признаков макропиносом, таких как размер, количество и местоположение. Описанный в настоящем описании способ особенно подходит для скрининга условий клеточной культуры, которые индуцируют макропиноцитоз, идентификации новых модуляторов или оптимизации концентраций лекарственного средства известных ингибиторов.
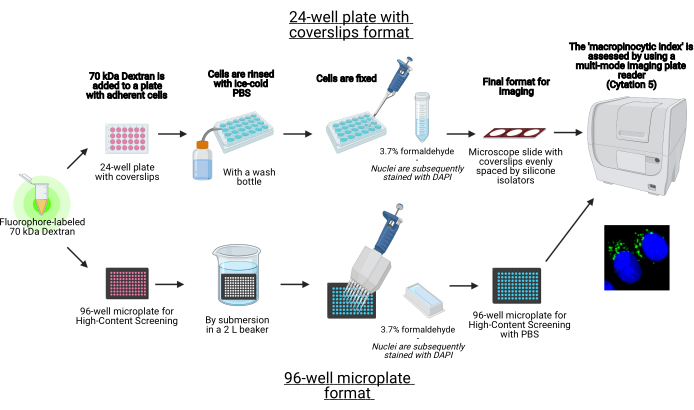
Рисунок 1: Схема автоматизированного анализа для определения «макропиноцитарного индекса» в адгезивных клетках. Создано с помощью BioRender. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
