Motor neuron (MN) diseases encompass a group of heterogenous conditions that include progressive degeneration leading to muscle wasting and paralysis as a primary clinical phenotype1. Although rare with a global prevalence of 4.5 per 100,000, this prevalence is expected to increase with an aging population2. Amyotrophic lateral sclerosis (ALS) is the most common MN disease (MND) and is typically fatal within a short time of diagnosis with no existing disease-modifying treatments available3. MNDs share in common a protracted presymptomatic phase with early molecular biomarker changes and functional imaging changes seen in patients4. Early presymptomatic cellular pathology is also observed in non-human disease models5,6,7,8. The study of early changes at the neuromuscular junction is important for understanding MN disease pathogenesis and may aid in developing early diagnostics and potential therapeutics.
A wealth of genetic and molecular tools exists in Drosophila to dissect the structure and function of the neuromuscular junction (NMJ, see9 for a review of the well-characterized larval NMJ). These tools combined with a short lifespan make Drosophila an excellent model to study neurodegenerative changes at the NMJ. Specifically, MNs innervating adult muscles are present throughout the ~90-day adult lifespan and are subject to normal aging processes10,11,12,13. The adult MNs therefore provide an opportunity to study slow degenerative changes in contrast to larval NMJs which exist for only a short ~1 week time-period prior to metamorphosis14,15.
Here, we describe a dissection procedure that allows us to conduct immunocytochemical analysis of MNs in the adult leg. Each adult leg is innervated by ~50 MNs, which synapse onto the associated leg muscular to drive locomotion. The leg anatomy, mechanical physiology, and neurobiology has been well described16,17,18. Axon arbors of leg MNs have previously been characterized by imaging through cuticle in back-filled or genetically labeled cell populations using the bipartite Gal4/UAS system and imaging methods have been published previously19. The dissection methods presented here preserves axon branching morphology and allows us to exploit a diverse range of antibodies to label different molecular components of the NMJ. Our previous work has focused on projections of a defined MN in the metathoracic (3rd) leg, which innervates the tibia levator muscle (tilm) and shows consistent arborization patterns and bouton numbers. Initially we studied age-dependent changes in Drosophila superoxide dismutase 1 (dsod1) mutants and found alterations consistent with dismantling of the NMJ20. These dissection methods offer the opportunity to better characterize slow degenerative changes at the NMJ for other ALS models, basic studies of aging and other MN associated diseases.
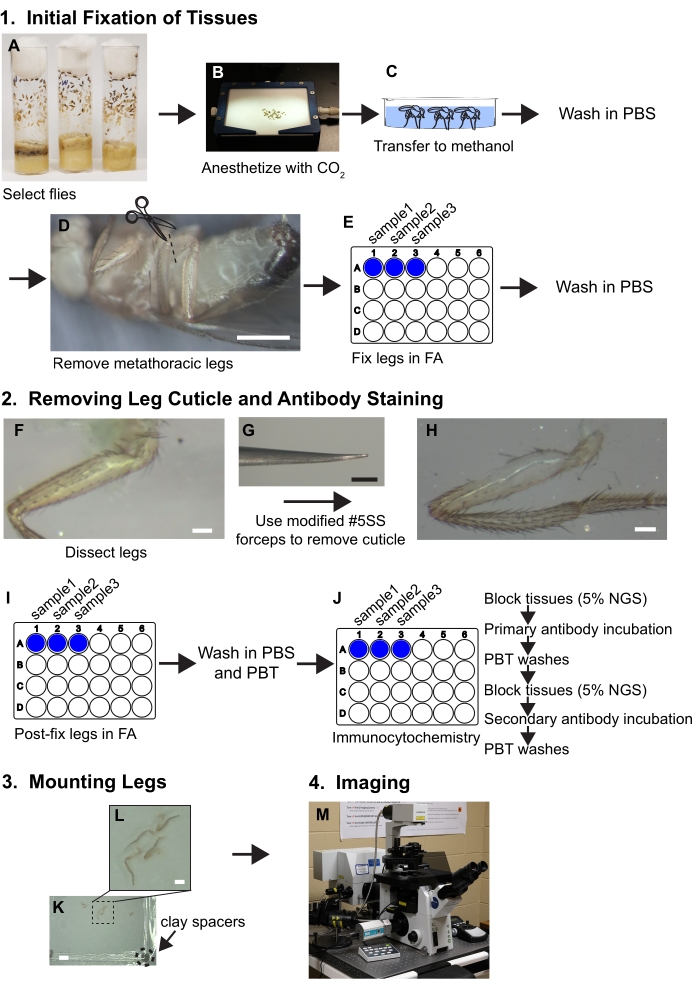
Figure 1. Workflow summary for dissecting legs. See protocol for detailed steps. (A,B) Flies are selected and anesthetized. (C) Flies are transferred to methanol and washed with PBS. (D) The metathoracic legs are removed at the base of the coxa while visualized with a dissecting microscope (~30x magnification); scale bar = 500 μm. (E) The legs are then fixed in 3.7% formaldehyde/PBS (FA) solution for 30 minutes within wells of 24-well plates and then FA is removed by washes with PBS. (F, G, H) Legs are transferred to silicone elastomer dissecting trays and a piece of cuticle is removed from the proximal femur using beveled forceps while visualized under a dissecting microscope at 80x; scale bar = 50 μm. (I) Legs are post-dissection fixed in FA and washed in PBS and then PBT (PBS+ 0.1% non-ionic surfactant). (J) Legs are subjected to immunocytochemical staining. (K,L) Legs are transferred to a glass slide, cleared in mounting media, and covered with a coverslip containing clay spacers; scale bars = 2 mm and 500 μm. (M) Legs are imaged by confocal microscopy. Please click here to view a larger version of this figure.
The procedures for preparing working solutions used in conjunction with this protocol are described in Table 1.
| Reagent | Preparation | Storage | |||
| PBS | Make a working stock of 1x PBS from a 10x PBS stock solution by diluting in distilled water. The pH of the working PBS stock should be 7.2- 7.4 | 4 °C for 1+ months until bacterial contamination is visible. | |||
| PBT | 1x PBS solution with 0.1% non-ionic surfactant. | 4 °C for 1+ months until bacterial contamination is visible. | |||
| FA | 3.7% formaldehyde solution made from a 37% formaldehyde stock and diluted in 1x PBS. | Room temperature. Make fresh each day of dissection | |||
| CAUTION: Formaldehyde supplied as 37% stock solution is a potential carcinogen and should be diluted in a fume hood. | |||||
| 5% NGS | 5% normal goat serum diluted in PBT. The serum used should match the species of the secondary antibody to be used. | 4 °C for several weeks until bacterial contamination is visible | |||
Table 1. Solutions for performing immunocytochemistry of the adult Drosophila leg.
1. Initial fixation of tissues
- Select approximately 10 flies for each genotype and age. Anesthetize on a fly pad under carbon dioxide (Figure 1A, B).
NOTE: Begin with more flies than necessary to ensure a large enough sample size after dissection. - Using a paintbrush, transfer flies to cold methanol in a glass well or dish for approximately 30 seconds to 1 minute (Figure 1C). The methanol solubilizes the cuticular hydrocarbons and flies can now be submerged rather than float in aqueous solutions.
- With forceps, carefully transfer flies to PBS. Rinse 3x in ice-cold PBS to remove excess methanol and keep flies in PBS on ice until dissection and fixation. At this point, dissect flies and fix within the shortest time possible (<30 minutes).
- To isolate legs, transfer flies to a silicone elastomer dissection dish filled with cold PBS, remove the metathoracic legs at the coxa using two pairs of #5 Dumont forceps or cut the legs with Vanna scissors (Figure 1D). Transfer the legs to a well in a plastic 24 well plate filled with 1 mL of PBS and keep the plate on ice until all legs are removed and transferred to wells.
NOTE: Each well can hold at least 20 legs. - Replace the PBS solution with 1 mL of FA solution and rotate on a nutator for 30 minutes (Figure 1E). The nutator setting should be set at a medium speed (17 rpm). Ensure the legs are completely submerged in FA solution during this time for adequate fixation.
- To remove FA solution, wash in 1 mL of PBS 3x quickly followed by an additional 3 washes for 5 minutes each in 1 mL of PBS. Hold tissues in 1 mL of PBS on ice prior to, and during the dissection steps described below.
2. Removing the Leg Cuticle and Antibody Staining
- Removing the leg cuticle
- The dissecting forceps are critical for success. Introduce slight parallel bends in both prongs at the end of #5 super fine forceps to provide a bevel that allows the cuticle to be grabbed superficially rather than be poked, which can ruin the tissue (Figure 1F, G).
NOTE: Prongs bent in parallel should still make contact with each other throughout the length of the prong when closed (Figure 2). - Transfer legs to a silicone elastomer dish in PBS for dissection. Orient a leg so that the anterior side is facing up (see Figure 3 for leg anatomy and orientation information). Using one pair of forceps, hold the tibia segment against the silicone elastomer dish. Using the other forceps held bevel side down, grab a piece of cuticle on the distal end of the femur and pull in the proximal direction toward the trochanter.
- Keep methodically removing cuticle until the naked muscle is visible throughout the proximal end of the femur (Figure 1F, G, H).
NOTE: Only make superficial contact with the legs using the beveled side of the forceps to avoid pulling muscle. - Once all legs are dissected, replace PBS with FA to post-fix legs for 30 minutes with shaking on a nutator at medium speed (Figure 1I). Wash samples in 1 mL of PBS for 3 times quick and then 3 times for 5 minutes each in PBT (Figure 1J).
NOTE: If staining with monoclonal antibody NC82 (anti-bruchpilot) to label active zones, post-fix for 20 minutes as this antigen is sensitive to longer fixations.
- The dissecting forceps are critical for success. Introduce slight parallel bends in both prongs at the end of #5 super fine forceps to provide a bevel that allows the cuticle to be grabbed superficially rather than be poked, which can ruin the tissue (Figure 1F, G).
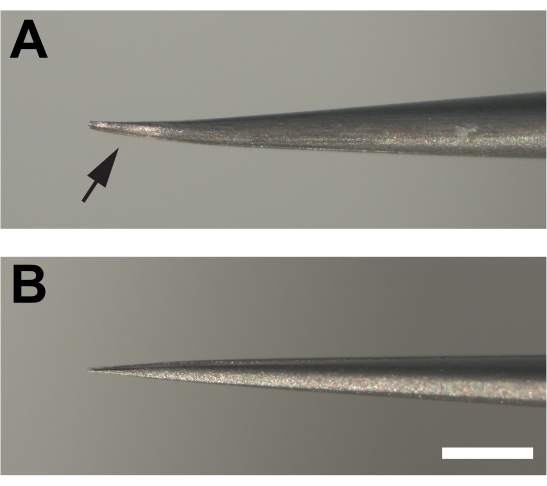
Figure 2. Modified forceps used for dissecting adult legs. (A) The ends of the forceps are bent and then flattened at the bottom (arrow) creating a bevel by filing on a sharpening stone. (B) In contrast, the prongs of unmodified forceps are not bent. Scale bar = 1 mm Please click here to view a larger version of this figure.
- Antibody staining
- To block tissues for antibody staining, replace 1 mL of PBT with blocking solution consisting of 1 mL of 5% NGS diluted in PBT. Incubate dissected legs for 4 hours at room-temperature or overnight at 4 °C while rocking on a nutator at medium speed (17 rpm). During all incubations, cover wells with sealing tape in addition to the plastic cover (Figure 1J).
NOTE: 24 well plates are used for immunocytochemistry rather than 1.5 mL or 2 mL microcentrifuge because previous attempts to use microcentrifuge tubes resulted in broken legs and damaged tissue. - Remove blocking solution and add 300 mL of primary antibodies diluted in fresh blocking solution. The small volume of antibody used should be sufficient to cover the tissues. Re-seal wells with laboratory sealing tape and plastic lid and incubate overnight at 4 °C with shaking on a nutator at medium speed (Figure 1J). The working antibody reagent information and concentrations used in for these studies are described in Table 2.
- Wash primary antibodies in 1 mL of PBT for 3 times briefly and then 3 times for 15 minutes each (Figure 1J).
NOTE: Diluted primary antibodies can be saved and reused if stored at 4 °C for up to 2 weeks. - Block tissues again in 1 mL of 5% NGS for at least 2 hours at room temperature or overnight at 4 C (Figure 1J).
- Remove the 5% NGS blocking solution and add 300 µL of the appropriate diluted fluorescent conjugated secondary antibodies. Additionally, add 1:2000 dilution of fluorescence-conjugated phalloidin to label muscle (Figure 1J).
- For secondary antibody incubations, seal wells with laboratory sealing tape and lid. Also, wrap plates in aluminum foil to protect fluorophores from light. Incubate for 6-8 hours at room-temperature or overnight at 4 °C.
- Wash secondary antibodies and phalloidin as described in step 2.2.3 above. Cover plates with aluminum foil in between washes to protect fluorophores from light.
- To block tissues for antibody staining, replace 1 mL of PBT with blocking solution consisting of 1 mL of 5% NGS diluted in PBT. Incubate dissected legs for 4 hours at room-temperature or overnight at 4 °C while rocking on a nutator at medium speed (17 rpm). During all incubations, cover wells with sealing tape in addition to the plastic cover (Figure 1J).
3. Mounting Legs
- Transfer legs to a slide using forceps and orient anterior side up. Cover the legs with mounting media (Figure 1K, L).
NOTE: The dissected legs can be aspirated with a P1000 pipette tip if the bottom bore is widened by cutting with a razor blade. - Add clay spacers to a 22×22 mm2 coverslip (#1.5 thickness) by scraping the coverslip corners across a small ball of modeling clay. Each corner should have a small amount of clay 1-2 mm thick (Figure 1K).
- To cover the slide, add the coverslip with the clay spacers facing towards the slide and carefully push on the corners until the coverslip just touches the surface of the femur.
- To prevent evaporation, seal the edges of the coverslip with nail polish and let dry in a dark place (about 10 minutes) before storing at 4 °C until imaging.
4. Imaging
- Image by confocal microscope (Figure 1M). Include a transmitted light channel to better assess the quality of the dissection and samples with visibly disrupted muscle fibers in the area of interest should be discarded.
- Begin imaging z-stacks with 20x magnification with 2x zoom and a total image depth of ~ 40 mm corresponding to the thickness of the femur. For fluorescent signal detection, capture images at resolutions consistent with Nyquist sampling (we are using 1024 x 1024 pixels with a dwell setting of 8-10 μs/pixel). The signal intensity should be in the linear range which is achieved by adjusting the high voltage gain settings. Once the gain settings are set for a series of samples within an experiment, they should not be altered so that signal intensities can be compared between samples.
- For imaging synaptic boutons and other subcellular structure, capture confocal images at ≥60x magnification. The detector settings should be in the linear range, while the pixel density, dwell settings and z-depth should be similar for images captured at lower magnification (step 4.1).
Figure 4 shows a representative example of a metathoracic leg stained with anti-hrp, anti-dlg, and phalloidin. For dissections that remove cuticle from the proximal portion of the femur, stereotyped arbors will be apparent near the tendon which is detected easily by autofluorescence. Note that antibody penetration into the leg occurs for a short distance beyond the region in which cuticle has been removed (Figure 4A). These regions can be imaged effectively when strong fluorescence signal is present. Imaging at low magnification (20x with a 2x zoom) allows an easy determination of 1) how much cuticle is removed, and 2) whether damage occurred during dissection. Increased magnification (60x) shows stereotyped projections onto the tilm (Figure 4B). Our work has focused on one MN, likely derived from the I- MN lineage which innervate the tilm in the proximal femur (box, Figure 4B and Figure 4C). Increasing magnification further (100x with 2x zoom) allows for effective visualization of synaptic boutons (Figure 4D).
To study morphological changes at the adult NMJ over time, we have previously used dsod1 mutants as a model of ALS. Bouton swelling occurs in aged dsod1H71Y mutants relative to dsod1null and dsod1+ (Figure 5A, B). At the larval NMJ, monoclonal antibody NC82 is often used to label active zones and this these structures can be easily visualized at the adult NMJ (Figure 5C). Weakly-positive HRP axon branches are abundant in dsod1H71Y mutants and these branches often show weak and diffuse BRP localization (arrows).
Important for neurodegeneration, commercially available antibodies can detect ubiquitinated proteins often found in aggregates, and we have detected ubiquitinated aggregates within terminal axons of MNs in aged mutant dsod1 flies as well as within muscle (Figure 6A). Also, antibodies labeling mitochondria can also detect morphological changes with age (Figure 6B).
For some preparations, muscle damage during the dissection makes the sample unusable. At first, such damage can be a common occurrence, but improvement occurs with practice. Examples of good dissections are show Figure 7A, C while poor dissections are shown in Figure 7B, D. Poor dissections cause disorganization of muscle fibers as detected by phase contrast microscopy (Figure 7B) or missing muscle fibers visualized by fluorescent-conjugated phalloidin (Figure 7D, arrow). The combination of using phalloidin to label muscle, and transmitted light images can help to detect such muscle damage which may not be apparent when viewing under a dissecting microscope.
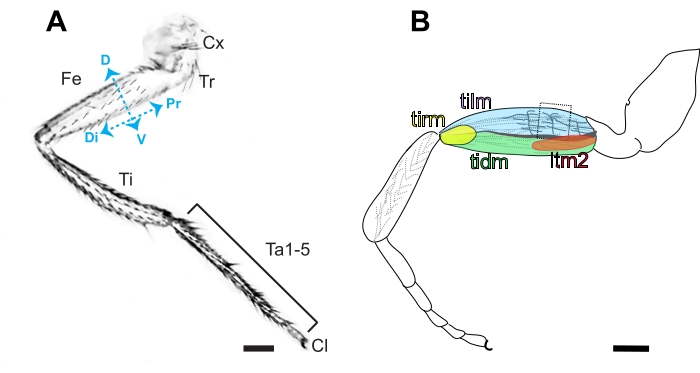
Figure 3. Anatomy of the Drosophila leg. (A) Diagram of the anterior side of a metathoracic leg, characterized by the presence of bristles in contrast to the prominent naked cuticle present on the posterior side. The segmented leg is comprised of the coxa (Cx), trochanter (Tr), femur (Fe), tibia (Ti), 5 tarsal segments (Ta1-5), and claw (Cl) in order from proximal (Pr) to distal (Di). Dorsal (D) and ventral (V) sides of the femur are also indicated. Scale bar =100 μm. (B) Diagram of the femur containing the tibia levator (tilm), tibia depressor (tidm), long tendon muscle 2 (ltm2) and tibia reductor (tirm) muscles and axon projections in the proximal femur innervating the tilm. These stereotyped axonal projections are presumed to be derived from I-lineage neuroblasts16, and the second arborization is the easiest to access by dissection (boxed). Scale bar = 100 μm. Please click here to view a larger version of this figure.
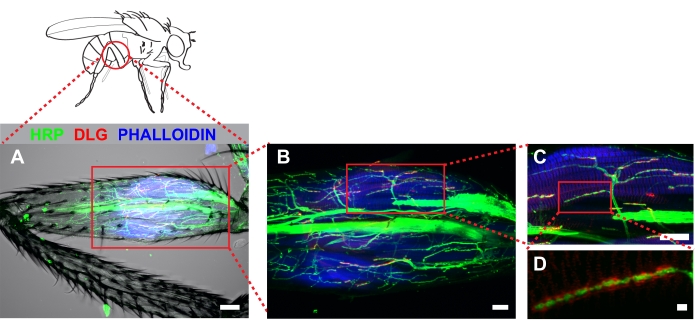
Figure 4. Dissected metathoracic femur revealed stereotyped MN architecture innervating the tilm. Immunocytochemistry was used to mark neurons (HRP), discs-large (DLG) and muscle (phalloidin). Z-stacks were captured by confocal microscopy imaging through the entire femur and shown as a maximum series projection. (A) A transmitted light channel was also included to illustrate no detectable muscle damage occurred during dissection. Image was captured at 20x magnification, scale bar =50 μm. (B) Identified arbors associated with the tilm (boxed area, center), scale bar =20 μm; and (C) at 60x magnification, scale bar = 20 μm. (D) DLG surrounding boutons was apparent in wild type animals when imaged at 100x magnification with 2x zoom; scale bar = 2 μm Please click here to view a larger version of this figure.
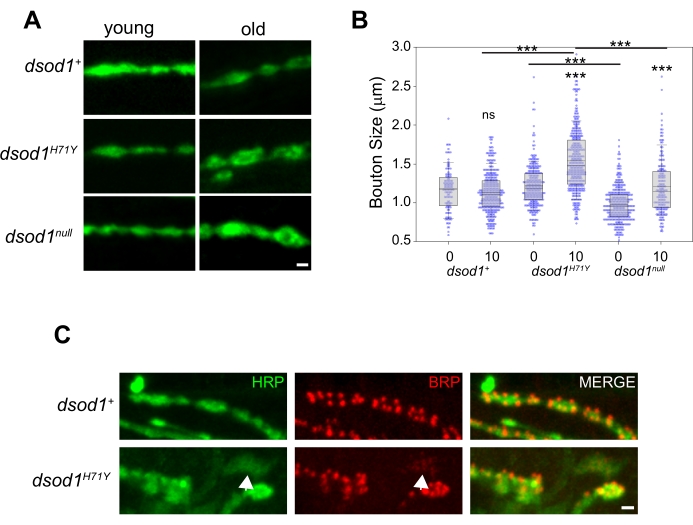
Figure 5. Example of age-dependent changes in bouton morphology which can be detected using the leg dissection technique. Bouton swelling is seen in aged dsod1H71Y mutants. (A) Representative images of HRP stained boutons from young (newly eclosed, day 0 adults) and old (day 10) flies, scale bar = 1 μm. (B) Bouton sizes were quantified from respective genotypes using the measure function within ImageJ. ***p<0.0001 (2-way ANOVA with post-hoc Tukey test). (C) Active zones within synaptic boutons labeled with monoclonal antibody NC82 (anti-bruchpilot); scale bar = 1 μm. Figure modified from20 Please click here to view a larger version of this figure.
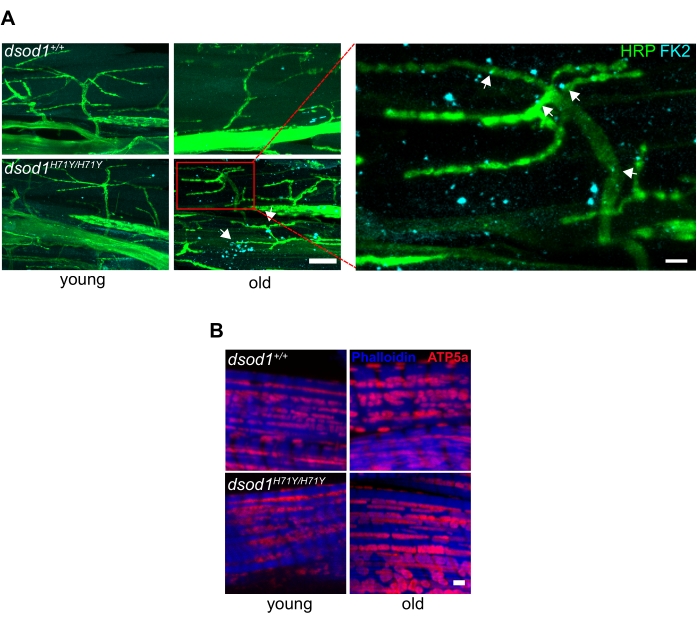
Figure 6. Common markers associated with neurodegenerative disease used in Drosophila leg preparations. (A) Subcellular polyubiquitinated aggregates are detected in terminal axons and muscle of aged dsod1H71Y flies. Immunocytochemistry using anti-polyubiquitin (FK2) detects puncta (arrows) in aged dsod1H71Y/H71Y, scale bar = 20 μm (left) and 5 μm (right). (B) Swollen mitochondria can be detected using anti-ATP5A, scale bar = 1 μm. Figure modified from20 Please click here to view a larger version of this figure.
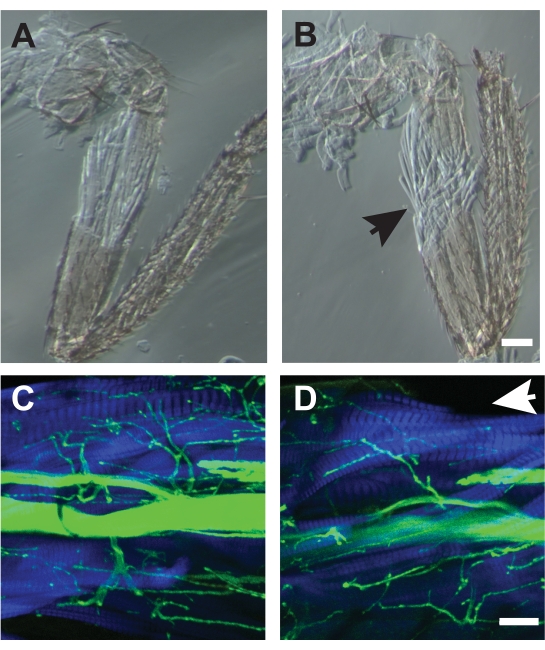
Figure 7. Examples of good and poor dissections. (A) Dissection which did not disrupt the underlying muscle architecture. (B) An example of a poor dissection which showed frayed muscle fibers (arrow) and could not be used for analysis. Both samples were imaged by phase-contrast microscopy. Scale bar = 50 μm. (C) Example of a good dissection imaged by confocal microscopy with HRP (green) and phalloidin (blue). (D) A poor dissection missing dorsal muscle fibers of the TILM missing (arrow). Scale bar = 50 μm Please click here to view a larger version of this figure.
| Antibody/Stain | Dilution* |
| Anti-ATP5A | 1:500 |
| Anti-bruchpilot (nc82) | 1:20 |
| Anti-cysteine string protein (ab49) | 1:50 |
| Anti-discs large (4F3) | 1:200 |
| Anti-hrp | 1:550 |
| Anti-polyubiquitin (FK2) | 1:1000 |
| Anti-repo (8D12) | 1:5 |
| Goat anti-mouse secondary antibody | 1:800 |
| Phalloidin | 1:2000 |
| *Dilute in 5% NGS |
Table 2. Antibodies and dilutions. The commercial suppliers of the antibodies used in these studies are listed in the Materials List.
