Selective Ribosome Profiling (SeRP) er den eneste metode til dato, der fanger og karakteriserer co-translationelle interaktioner, in vivo, på en direkte måde 1,2,3,4,5,6. SeRP muliggør global profilering af interaktioner af enhver faktor med oversættelse af ribosomer i nærkodonopløsning 2,7.
Metoden er afhængig af flashfrysning af voksende celler og bevarelse af aktiv translation. Cellelysater behandles derefter med RNase I for at fordøje alt mRNA i cellen undtagen ribosombeskyttede mRNA-fragmenter kaldet “ribosomfodspor”. Prøven opdeles derefter i to dele; en del bruges direkte til isolering af alle de cellulære ribosomale fodspor, der repræsenterer al løbende translation i cellen. Den anden del anvendes til affinitetsrensning af den specifikke delmængde af ribosomer forbundet med en faktor af interesse, for eksempel: modificerende enzymer, translokationsfaktorer, foldning af chaperoner og komplekse samlingsinteraktioner. De affinitetsrensede ribosomale fodspor kaldes samlet interactomet. Derefter ekstraheres de ribosombeskyttede mRNA’er og bruges til cDNA-biblioteksgenerering efterfulgt af dyb sekventering.
Sammenlignende analyse af de samlede translatom- og interactomprøver muliggør identifikation af alle orfs, der er forbundet med interessefaktoren, samt karakterisering af hver orf-interaktionsprofil. Denne profil rapporterer de præcise engagementsstart- og afslutningssekvenser, hvorfra man kan udlede de afkodede kodoner og de respektive rester af den nye polypeptidkæde samt på ribosomhastighedsvariationerne under interaktionen 7,8. Figur 1 viser protokollen som en skematisk.
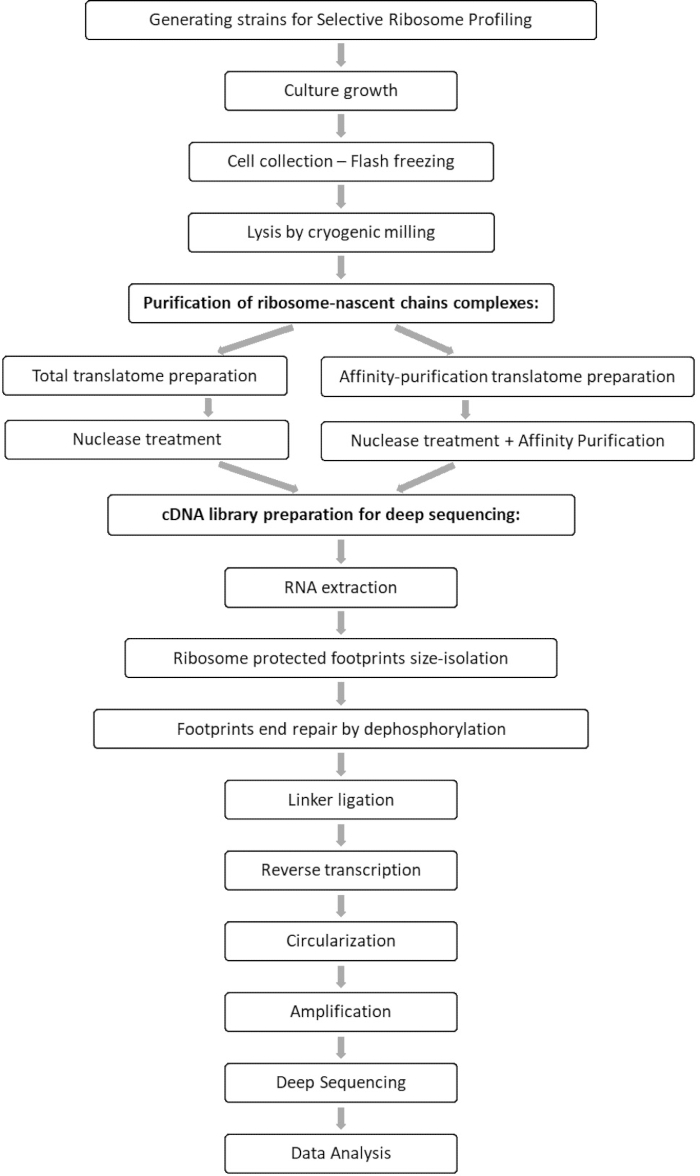
Figur 1: En oversigt over SeRP-protokollen. Denne protokol kan udføres i sin helhed inden for 7-10 dage. Klik her for at se en større version af denne figur.
