Selective Ribosome Profiling (SeRP) is tot op heden de enige methode die co-translationele interacties, in vivo, op een directe maniervastlegt en karakteriseert 1,2,3,4,5,6. SeRP maakt globale profilering van interacties van elke factor mogelijk met het vertalen van ribosomen in bijna codonresolutie 2,7.
De methode is gebaseerd op het flashvriezen van groeiende cellen en het behoud van actieve translatie. Cellysaten worden vervolgens behandeld met RNase I om al het mRNA in de cel te verteren, behalve ribosoom-beschermde mRNA-fragmenten die “ribosoomvoetafdrukken” worden genoemd. Het monster wordt vervolgens in twee delen gesplitst; een deel wordt direct gebruikt voor de isolatie van alle cellulaire ribosomale voetafdrukken, die alle lopende vertaling in de cel vertegenwoordigen. Het tweede deel wordt gebruikt voor de affiniteitszuivering van de specifieke subset van ribosomen geassocieerd met een factor van belang, bijvoorbeeld: het modificeren van enzymen, translocatiefactoren, vouw chaperonnes en complex-assemblage interacties. De affiniteitsgezuiverde ribosomale voetafdrukken worden gezamenlijk het interactoom genoemd. Vervolgens worden de ribosoombeveiligde mRNA’s geëxtraheerd en gebruikt voor het genereren van cDNA-bibliotheken, gevolgd door diepe sequencing.
Vergelijkende analyse van de totale translatoom- en interactoommonsters maakt het mogelijk om alle orfs te identificeren die verband houden met de factor van belang, evenals karakterisering van elk orf-interactieprofiel. Dit profiel rapporteert de precieze betrokkenheids- en beëindigingssequenties waaruit men de gedecodeerde codons en de respectieve residuen van de opkomende polypeptideketen kan afleiden, evenals op de ribosoomsnelheidsvariaties tijdens de interactie 7,8. Figuur 1 geeft het protocol weer als een schema.
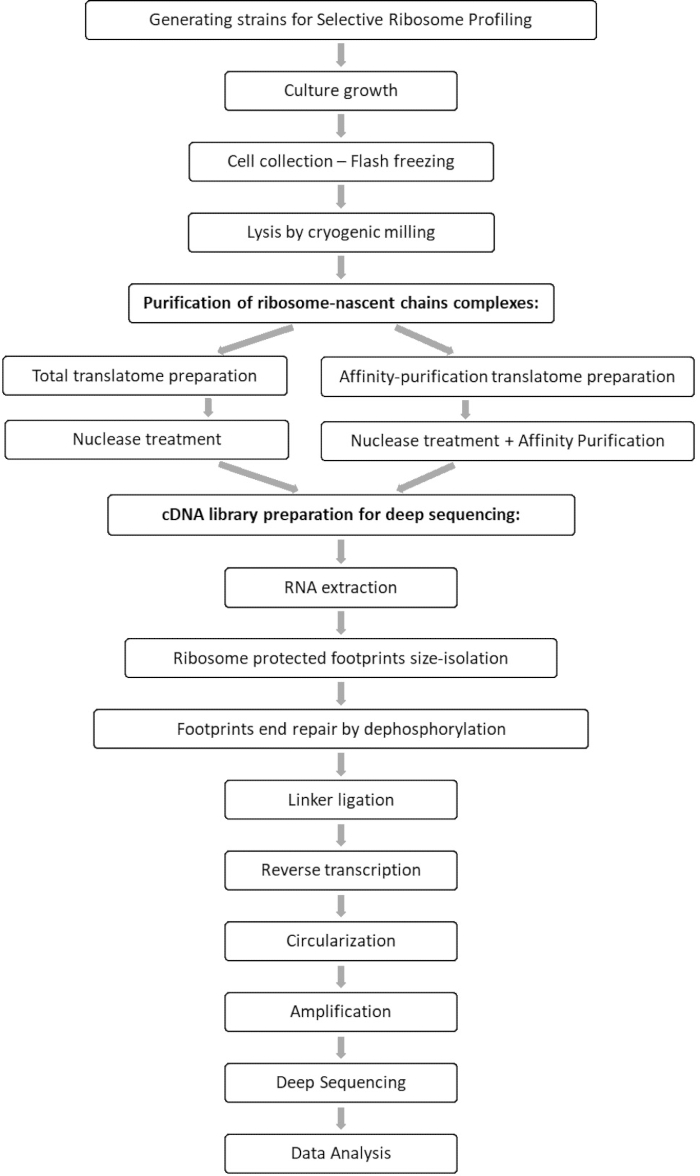
Figuur 1: Een overzicht van het SeRP-protocol. Dit protocol kan in zijn geheel binnen 7-10 dagen worden uitgevoerd. Klik hier om een grotere versie van deze figuur te bekijken.
