Lerofilage de l’ibosome RR (SeRP) est la seule méthode, à ce jour, qui capture et caractérise les interactions co-translationnelles, in vivo, de manière directe 1,2,3,4,5,6. SeRP permet un profilage global des interactions de n’importe quel facteur avec la traduction des ribosomes en résolution proche du codon 2,7.
La méthode repose sur la congélation éclair des cellules en croissance et la préservation de la traduction active. Les lysats cellulaires sont ensuite traités avec de la RNase I pour digérer tout l’ARNm dans la cellule, à l’exception des fragments d’ARNm protégés par les ribosomes appelés « empreintes de ribosomes ». L’échantillon est ensuite divisé en deux parties; une partie est directement utilisée pour l’isolement de toutes les empreintes ribosomiques cellulaires, représentant toute la traduction en cours dans la cellule. La deuxième partie est utilisée pour la purification d’affinité du sous-ensemble spécifique de ribosomes associés à un facteur d’intérêt, par exemple: enzymes modificatrices, facteurs de translocation, chaperons de repliement et interactions d’assemblage complexe. Les empreintes ribosomiques purifiées par affinité sont collectivement appelées interactome. Ensuite, les ARNm protégés par les ribosomes sont extraits et utilisés pour la génération de bibliothèque d’ADNc, suivie d’un séquençage en profondeur.
L’analyse comparative des échantillons totaux de translatome et d’interactome permet d’identifier tous les orfs associés au facteur d’intérêt, ainsi que la caractérisation de chaque profil d’interaction orf. Ce profil rapporte les séquences précises d’apparition et de terminaison de l’engagement à partir desquelles on peut déduire les codons décodés et les résidus respectifs de la chaîne polypeptidique émergente, ainsi que sur les variations de vitesse des ribosomes au cours de l’interaction 7,8. La figure 1 illustre le protocole sous forme de schéma.
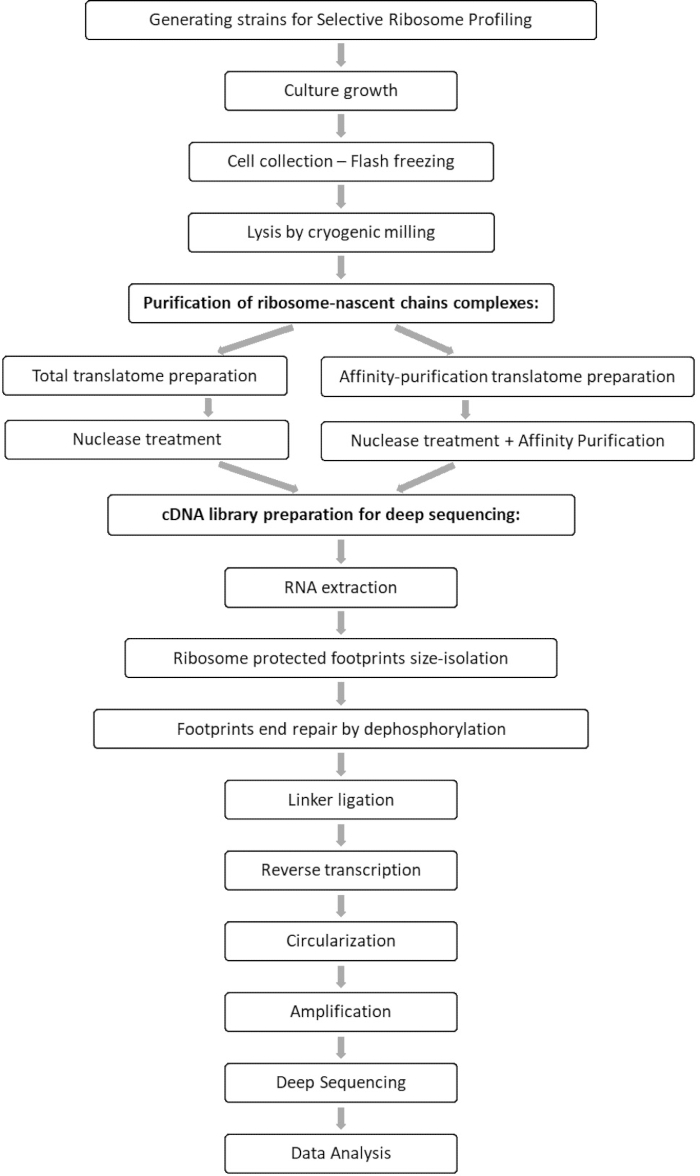
Figure 1 : Vue d’ensemble du protocole SeRP. Ce protocole peut être effectué dans son intégralité dans un délai de 7 à 10 jours. Veuillez cliquer ici pour voir une version agrandie de cette figure.
