Selective Ribosome Profiling (SeRP) היא השיטה היחידה, עד כה, הלוכדת ומאפיינת אינטראקציות תרגום משותף, in vivo, באופן ישיר 1,2,3,4,5,6. SeRP מאפשר פרופיל גלובלי של אינטראקציות של כל גורם עם תרגום ריבוזומים ברזולוציה של כמעט קודון 2,7.
השיטה מסתמכת על הקפאת הבזק של תאים גדלים ושמירה על תרגום פעיל. לאחר מכן, ליזטים של תאים מטופלים באמצעות RNase I כדי לעכל את כל ה-mRNA בתא, למעט שברי mRNA המוגנים על-ידי ריבוזומים המכונים “עקבות ריבוזומים”. לאחר מכן מתפצלים המדגם לשני חלקים; חלק אחד משמש ישירות לבידוד של כל עקבות הריבוזומליות התאיות, המייצגות את כל התרגום המתמשך בתא. החלק השני משמש לטיהור זיקה של תת-קבוצה ספציפית של ריבוזומים הקשורים לגורם עניין, לדוגמה: שינוי אנזימים, גורמי טרנסלוקציה, מלווים מתקפלים ואינטראקציות של הרכבה מורכבת. עקבות הריבוזומליות המטוהרות זיקה נקראות באופן קולקטיבי האינטראקציה. לאחר מכן, ה-mRNA המוגנים בריבוזום מופקים ומשמשים ליצירת ספריית cDNA, ולאחר מכן לריצוף עמוק.
ניתוח השוואתי של סך כל דגימות התרגום והאינטראקציה מאפשר זיהוי של כל האורפים המתקשרים עם גורם העניין, כמו גם אפיון של כל פרופיל אינטראקציה של orf. פרופיל זה מדווח על רצפי ההתחלה והסיום המדויקים של ההתקשרות שמהם ניתן להסיק את הקודונים המפוענחים ואת השאריות המתאימות של שרשרת הפוליפפטידים המתהווה, כמו גם על שינויי מהירות הריבוזומים במהלך האינטראקציה 7,8. איור 1 מתאר את הפרוטוקול כשרטוט.
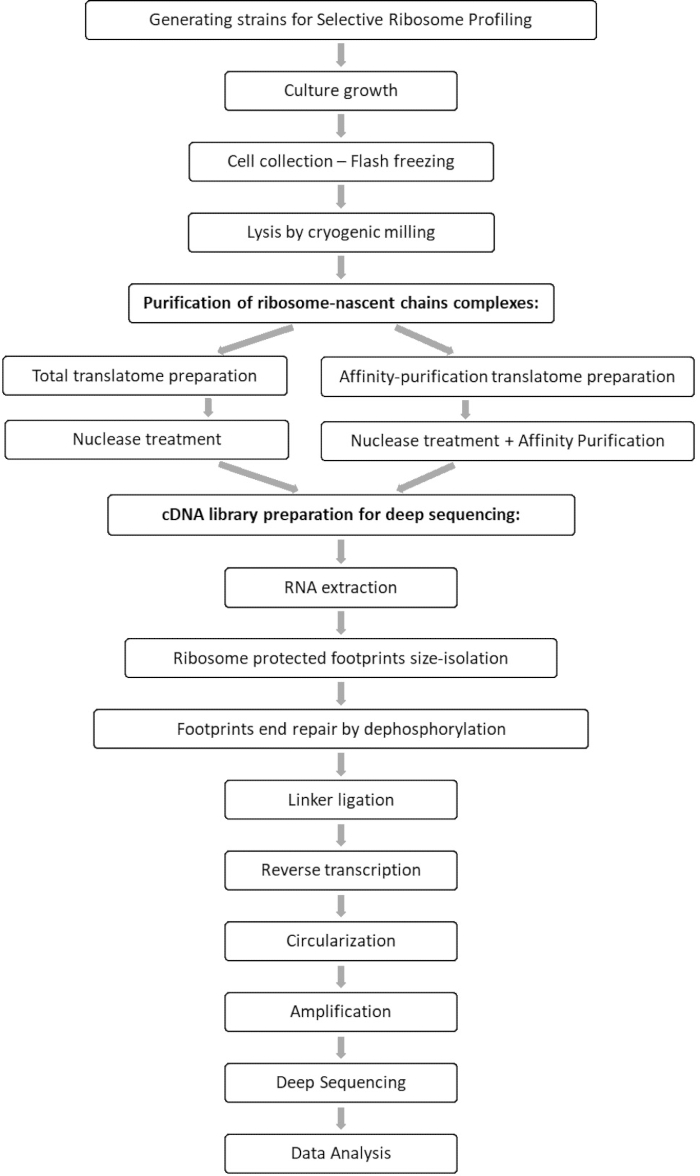
איור 1: סקירה כללית של פרוטוקול SeRP. פרוטוקול זה יכול להתבצע בשלמותו תוך 7-10 ימים. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
