Selective Ribosome Profiling (SeRP) é o único método, até o momento, que captura e caracteriza interações co-translacionais, in vivo, de forma direta 1,2,3,4,5,6. O SeRP permite o perfil global de interações de qualquer fator com a tradução de ribossomos na resolução 2,7 de códon próximo.
O método se baseia no congelamento flash de células em crescimento e na preservação da tradução ativa. Os lises celulares são então tratados com RNase I para digerir todo o mRNA na célula, exceto fragmentos de mRNA protegidos por ribossomos denominados “pegadas ribossósas”. A amostra é então dividida em duas partes; uma parte é diretamente usada para o isolamento de todas as pegadas ribossômicas celulares, representando toda a tradução contínua na célula. A segunda parte é utilizada para a afinidade-purificação do subconjunto específico de ribossomos associados a um fator de interesse, por exemplo: modificação de enzimas, fatores de translocação, acompanhantes dobráveis e interações de montagem complexa. As pegadas ribossômicas purificadas por afinidade são coletivamente chamadas de interactome. Em seguida, os mRNAs protegidos por ribossomo são extraídos e usados para a geração de biblioteca cDNA, seguido de sequenciamento profundo.
A análise comparativa das amostras totais de translatome e interactome permite a identificação de todos os orfs que associam ao fator de interesse, bem como a caracterização de cada perfil de interação orf. Este perfil relata as sequências precisas de início e término de engajamento a partir das quais se pode inferir os códons decodificados e os respectivos resíduos da cadeia emergente de polipeptídeos, bem como sobre as variações de velocidade ribossosome durante a interação 7,8. A Figura 1 retrata o protocolo como um esquema.
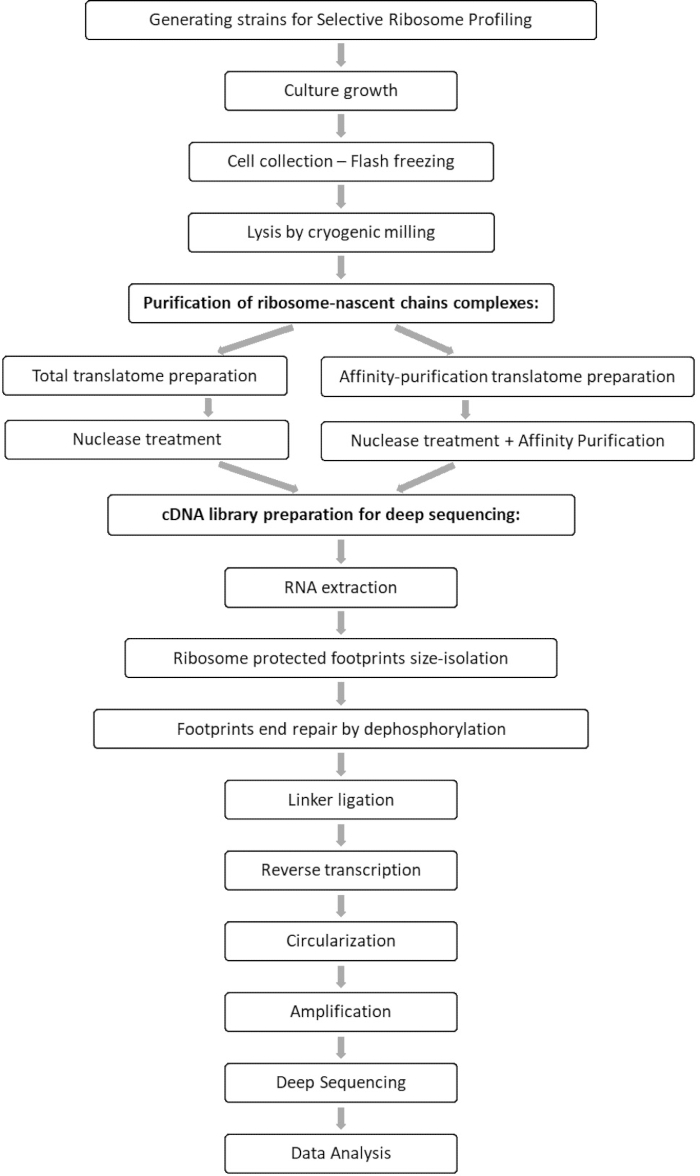
Figura 1: Uma visão geral do protocolo SeRP. Este protocolo pode ser realizado em sua totalidade dentro de 7-10 dias. Clique aqui para ver uma versão maior desta figura.
