Elrofiling Ribosome P(SeRP) es el único método, hasta la fecha, que captura y caracteriza las interacciones co-traslacionales, in vivo, de manera directa 1,2,3,4,5,6. SeRP permite el perfil global de las interacciones de cualquier factor con la traducción de ribosomas en resolución de codón cercano 2,7.
El método se basa en la congelación instantánea de las células en crecimiento y la preservación de la traducción activa. Los lisados celulares se tratan con RNasa I para digerir todo el ARNm en la célula, excepto los fragmentos de ARNm protegidos por ribosomas denominados “huellas de ribosomas”. La muestra se divide entonces en dos partes; una parte se utiliza directamente para el aislamiento de todas las huellas ribosómicas celulares, que representan toda la traducción en curso en la célula. La segunda parte se utiliza para la afinidad-purificación del subconjunto específico de ribosomas asociados con un factor de interés, por ejemplo: enzimas modificadoras, factores de translocación, chaperonas plegables e interacciones complejo-ensamblaje. Las huellas ribosómicas purificadas por afinidad se denominan colectivamente el interactoma. Luego, los ARNm protegidos por ribosomas se extraen y se utilizan para la generación de bibliotecas de ADNc, seguidos de una secuenciación profunda.
El análisis comparativo de las muestras totales de translatoma e interactoma permite la identificación de todos los orfs que se asocian con el factor de interés, así como la caracterización de cada perfil de interacción orf. Este perfil informa las secuencias precisas de inicio y terminación del compromiso a partir de las cuales se pueden inferir los codones decodificados y los residuos respectivos de la cadena polipeptídica emergente, así como sobre las variaciones de velocidad del ribosoma durante la interacción 7,8. La figura 1 muestra el protocolo como un esquema.
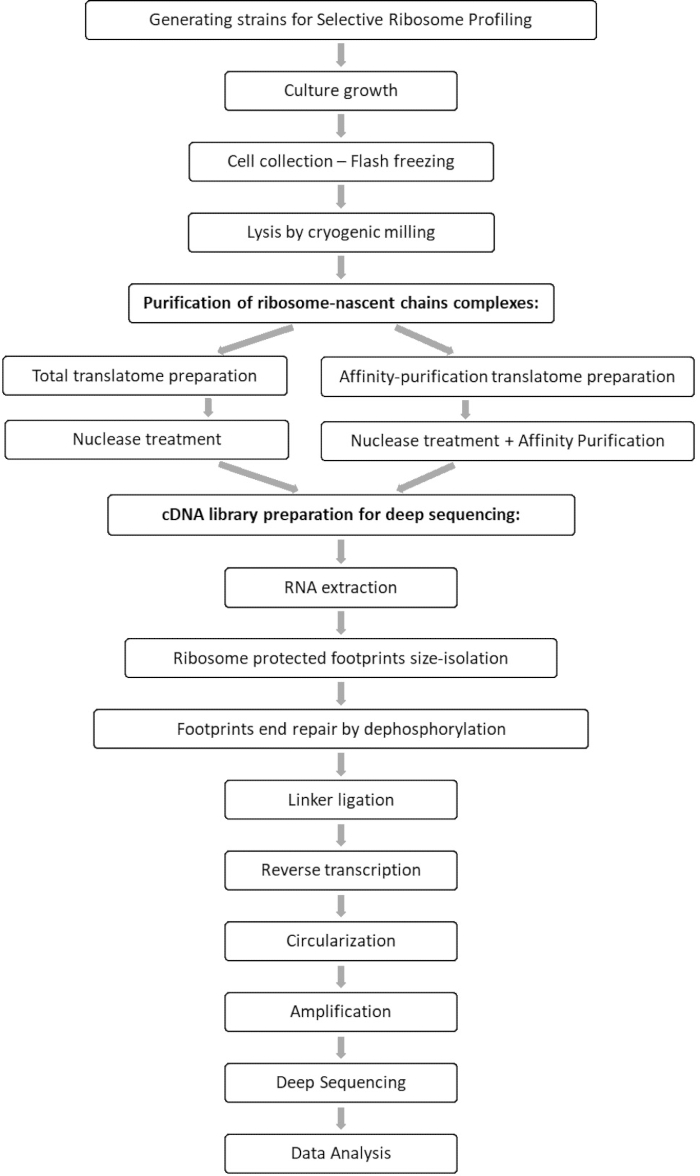
Figura 1: Una visión general del protocolo SeRP. Este protocolo se puede realizar en su totalidad en un plazo de 7-10 días. Haga clic aquí para ver una versión más grande de esta figura.
