Selective Ribozom Profiling (SeRP), bugüne kadar, ko-translasyonel etkileşimleri, in vivo, doğrudan bir şekilde 1,2,3,4,5,6 olarak yakalayan ve karakterize eden tek yöntemdir. SeRP, ribozomların 2,7’ye yakın çözünürlükte çevrilmesiyle herhangi bir faktörün etkileşimlerinin global profillenmesini sağlar.
Yöntem, büyüyen hücrelerin flaş dondurulmasına ve aktif çevirinin korunmasına dayanır. Hücre lizatları daha sonra “ribozom ayak izleri” olarak adlandırılan ribozom korumalı mRNA fragmanları hariç hücredeki tüm mRNA’yı sindirmek için RNaz I ile muamele edilir. Numune daha sonra iki bölüme ayrılır; Bir kısım doğrudan tüm hücresel ribozomal ayak izlerinin izolasyonu için kullanılır ve hücrede devam eden tüm translasyonu temsil eder. İkinci bölüm, bir ilgi faktörü ile ilişkili ribozomların spesifik alt kümesinin afinite saflaştırılması için kullanılır, örneğin: enzimleri değiştirmek, translokasyon faktörleri, şaperonları katlamak ve karmaşık montaj etkileşimleri. Afinite saflaştırılmış ribozomal ayak izleri topluca interaktom olarak adlandırılır. Daha sonra, ribozom korumalı mRNA’lar ekstrakte edilir ve cDNA kütüphanesi üretimi için kullanılır, ardından derin dizileme yapılır.
Toplam translatom ve interaktom örneklerinin karşılaştırmalı analizi, ilgi faktörü ile ilişkili tüm orfların tanımlanmasına ve her bir orf etkileşim profilinin karakterizasyonuna izin verir. Bu profil, kodu çözülmüş kodonları ve ortaya çıkan polipeptit zincirinin ilgili kalıntılarını ve ayrıca etkileşim 7,8 sırasındaki ribozom hız değişimlerini çıkarabileceğiniz kesin katılım başlangıcı ve sonlandırma dizilerini rapor eder. Şekil 1’de protokol şematik olarak gösterilmektedir.
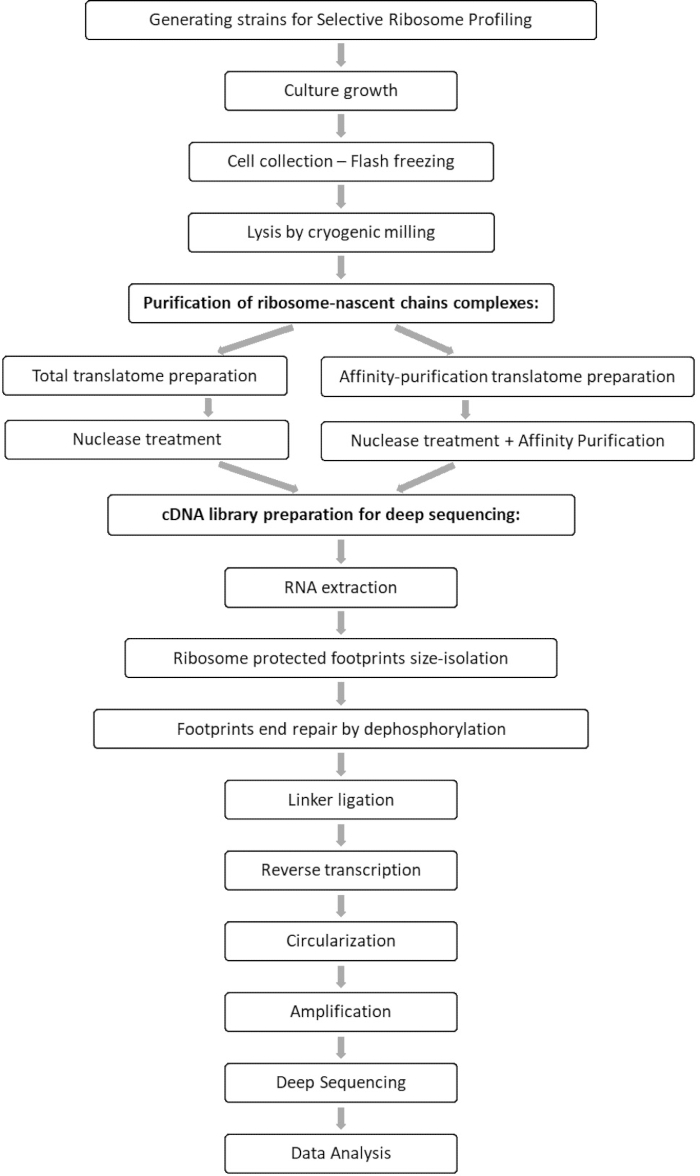
Şekil 1: SeRP protokolüne genel bakış. Bu protokol 7-10 gün içinde bütünüyle gerçekleştirilebilir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
