Selective Ribosome Profiling (SeRP) er den eneste metoden, til dags dato, som fanger opp og karakteriserer co-translational interaksjoner, in vivo, på en direkte måte 1,2,3,4,5,6. SeRP muliggjør global profilering av interaksjoner av enhver faktor med oversettelse av ribosomer i nær codon-oppløsning 2,7.
Metoden er avhengig av flashfrysing av voksende celler og bevaring av aktiv oversettelse. Cellelysater behandles deretter med RNase I for å fordøye all mRNA i cellen bortsett fra ribosombeskyttede mRNA-fragmenter kalt “ribosome fotavtrykk”. Prøven deles deretter i to deler. en del brukes direkte til isolering av alle de cellulære ribosomale fotavtrykkene, som representerer alle pågående oversettelser i cellen. Den andre delen brukes til affinitetsrensing av den spesifikke undergruppen av ribosomer forbundet med en interessefaktor, for eksempel: modifisere enzymer, translokasjonsfaktorer, sammenleggbare anstander og komplekse monteringsinteraksjoner. De affinitetsrensede ribosomale fotavtrykkene betegnes samlet som interaktivitet. Deretter ekstraheres ribosombeskyttede mRNAer og brukes til cDNA-bibliotekgenerering, etterfulgt av dyp sekvensering.
Komparativ analyse av de totale translatom- og interactome-prøvene gjør det mulig å identifisere alle orfer som er knyttet til interessefaktoren, samt karakterisering av hver orf interaksjonsprofil. Denne profilen rapporterer de nøyaktige engasjements- og avslutningssekvensene som man kan utlede de dekodede codonene og de respektive rester av den fremvoksende polypeptidkjeden, samt på ribosomets hastighetsvariasjoner under samspillet 7,8. Figur 1 viser protokollen som et skjema.
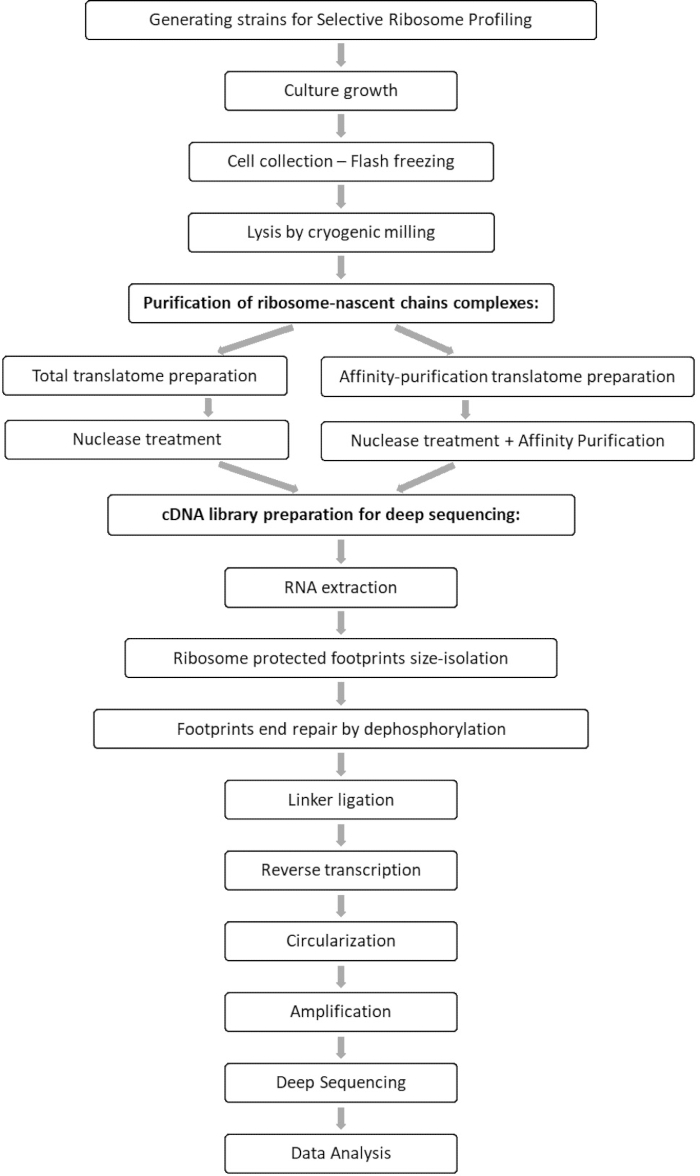
Figur 1: En oversikt over SeRP-protokollen. Denne protokollen kan utføres i sin helhet innen 7-10 dager. Klikk her for å se en større versjon av denne figuren.
