מתילציה של דנ”א היא סימן אפיגנטי חשוב המווסת את ביטוי הגנים ואת מבנה הכרומטין. מתילציה מתרחשת בעיקר בדינוקלאוטידים של CpG — ציטוזין ואחריו גואנוזין; קבוצת המתיל מתווספת למיקום 5 של ציטוזין. יש צורך בתבניות מתילציה נכונות של DNA, ובכך בביטוי גנים תקין, להתפתחות ותפקוד מתאימים של התאים. מצבי מחלה רבים נקשרו לשינויים בתבנית המתילציה הרגילה 1,2,3. לדוגמה, קיים קשר בין התחלת סרטן והתקדמותו לבין שינויים בתבנית המתילציה של הדנ”א. בדרך כלל, תאים סרטניים מציגים רמות כלליות נמוכות יותר של מתילציטוזין, אשר תורם לחוסר יציבות הגנום. יחד עם זאת, המתילציטוזין שנמצא בגנום מרוכז באזורים המקדמים של גנים מדכאי גידול, מה שמוביל להשתקת גנים של חלבונים חשובים אלה. יש לציין כי שינויים אפיגנטיים הם דינמיים והפיכים, בניגוד למוטציות הדנ”א הקשורות לגידול. זה הפך את החלבונים המעורבים בוויסות גנים אפיגנטיים למטרות מעניינות של תרופות 2,4.
מתיל-טרנספראזות דנ”א (DNMTs) הם החלבונים האחראים על ייצור ושמירה על תבניות מתילציה של דנ”א. שלושה איזוזימים פעילים קטליטיים, DNMT1, DNMT3a ו-DNMT3b, קיימים בבני אדם. במהלך הפיתוח וההתמיינות קובעים המתיל-טרנספראזות של דה נובו, DNMT3a ו-DNMT3b, תבניות מתילציה. שני האנזימים יכולים לקשור את חלבון ה-DNMT3L שאינו פעיל באופן קטליטי ליצירת קומפלקסים המציגים פעילות מוגברת 1,5. לאחר חלוקת התא, תאי הבת מכילים דנ”א שעבר המימתילציה – דנ”א המכיל מתילציטוזין בגדיל אחד בלבד של הדופלקס – מאחר שהדנ”א המסונתז החדש נטול סימני מתילציה. תפקידו העיקרי של DNMT1 הוא לבצע מתילציה של דנ”א המימתילציה זה, ובכך לבסס מחדש את תבנית המתילציה המלאה 1,5.
הקשרים בין פעילות DNMT לסרטן מבוססים היטב. ביטוי יתר של DNMT1, בין אם על ידי מנגנוני שעתוק או לאחר תרגום, הוא תוצאה של מספר מסלולים אונקוגניים נפוצים 6,7,8,9. גישות גנטיות להפחתת פעילות DNMT1 באמצעות אללים היפומורפיים גורמות לירידה בהיווצרות הגידול בעכברי Apc(Min) 10. אוליגונוקלאוטידים אנטיסנסים שמפילים DNMT1 מעכבים ניאופלזיה בתרבית תאים ובמודלים של גידולי עכברים11,12. לפיכך, עיכוב פעילות DNMT1 נראה כמו גישה מבטיחה לטיפול בסרטן. עם זאת, התפקידים שהאיזוזימים של DNMT3 ממלאים אינם כה פשוטים. מוטציות DNMT3a נמצאות בלוקמיה מיאלואידית חריפה13 ובתסמונת מיאלודיספלסטית14. לפחות אחת מהמוטציות שזוהו הוכחה כמפחיתה את פעילות המתילציה של הדנ”א של האנזים15. עם זאת, DNMT3b מתבטא יתר על המידה בסרטן השד16 ובסרטן המעי הגס17. כאשר איזוזימים שונים של DNMT ממלאים תפקידים שונים בסרטן, זיהוי מעכבים ספציפיים לאיזוזים יהיה קריטי. לא רק שתרכובות אלה יהיו שימושיות לפיתוח טיפולים, אלא שמעכבים ספציפיים לאיזוזים יהיו גם כלי רב ערך לנתח את התפקיד של כל איזוזימה DNMT באטיולוגיה של סרטן.
מספר מעכבי DNMT דווחו בספרות. מעכבי DNMT ידועים ניתן לחלק לשני סוגים: נוקלאוזיד ולא נוקלאוזיד. מעכבי נוקלאוזיד הם בדרך כלל אנלוגים של ציטידין. תרכובות אלה משולבות בדנ”א ולוכדות באופן קוולנטי DNMTs. 5-azacytidine ו-5-aza-2′-deoxycytidine אושרו לטיפול בתסמונת מיאלודיספלסטית ולוקמיה מיאלואידית חריפה 4,18. הרעילות הגבוהה, הזמינות הביולוגית הנמוכה וחוסר היציבות הכימית של תרכובות אלה מהווים בעיות. עבודה שוטפת בוחנת את היעילות של הדור הבא של מעכבי נוקלאוזידים; SGI-110, המופק מ-5-aza-2′-deoxycytidine, הוא דוגמה אחת19,20. מעכבי נוקלאוזידים אינם ספציפיים לאיזוזימים וישביתו כל איזוזימה DNMT שנתקלים בה. לכן, טיפול בחומר מתיל של נוקלאוזיד גורם לדלדול כל האיזוזימים של DNMT 4,18. מעכבים שאינם נוקלאוזידים אינם צריכים להיות משולבים בדנ”א כדי להפעיל את ההשפעות המעכבות שלהם. במקום זאת, מולקולות אלה נקשרות ישירות ל-DNMTs, מה שמציג את האפשרות לעיכוב ספציפי לאיזוזימים. מספר מעכבי נוקליאוזיד התגלו עד כה, כולל SGI-1027 21, הידרלזין 22, פרוקאינאמיד 23, RG108 ונגזרותיו24, ומוצרים טבעיים, (−)-אפיגלוקטכין 3-גלאט (EGCG)25 וחומצה לאקאית A 26,27. רוב המעכבים שאינם נוקלאוזידים שהתגלו עד כה אינם איזוזימים סלקטיביים או מציגים העדפות חלשות לאיזוזימה אחת של DNMT. בנוסף, יש לשפר את העוצמה של מולקולות אלה, במיוחד בתאים 4,18. לכן, יש צורך לגלות או לפתח מעכבי DNMT חזקים יותר, איזוזימים-סלקטיביים.
מכשול לגילוי מעכבי מולקולות קטנות חדשות של DNMTs הוא המבחנים המייגעים המשמשים באופן מסורתי לבחינת פעילות DNMT28. מבחנים הם בדרך כלל רציפים עם מספר שלבים. הפעילות האנזימטית של DNMTs עדיין נבדקת באופן שגרתי באמצעות S-אדנוזיל מתיונין רדיואקטיבי (SAM)29,30,31,32,33,34. כמו כן פותחו בדיקות לא רדיואקטיביות למתילציה של DNA. לדוגמה, בדיקות המשתמשות באנדונוקלאזות הגבלה רגישות למתיל ובאלקטרופורזה כדי להפריד בין תוצרי העיכול תוארו35,36. סוגים אלה של מבחנים רציפים, מרובי שלבים, אינם ניתנים בקלות לגילוי תרופות. מאז אמצע שנות ה-2000 פותחו מספר מבחני מתילציה של דנ”א עם תפוקה גבוהה יותר28. בדיקת קרבה של סינטילציה שימשה לבדיקת מעכבי DNMT137. בדיקה נוספת המשתמשת באנדונוקלאז הגבלה רגיש למתיל שימשה לבדיקת מעכבי DNMT3a25,38. בעוד ששני המבחנים אפשרו תפוקה גבוהה יותר מאשר מבחני מתילציה מסורתיים של DNA, הבדיקות דורשות שלבים מרובים ואינן מאפשרות תצפית על פעילות מתילציה בזמן אמת. לאחרונה תוארה בדיקת קינטיקה רציפה המשלבת את היווצרותו של S-אדנוזילהומוציסטאין (SAH), תוצר אחד של תגובת המתילציה, לשינוי הספקטרוסקופי ב-340 ננומטר הקשור לחמצון NADPH39. בדיקה זו משתמשת בשלושה אנזימי צימוד כדי ליצור אות ספקטרוסקופי.
פיתחנו בדיקת מתילציה של דנ”א מצומד אנדונוקלאז המבוססת על פלואורסצנציה, המשתמשת באנזים צימוד יחיד הזמין מסחרית ויכולה להפיק נתונים בזמן אמת (איור 1). אוליגונוקלאוטיד סיכת שיער המכיל שלושה מתילציטוסינים משמש כמצע. דנ”א המצע מכיל פלואורופור בקצה 5′ וקוונצ’ר בקצה 3′. מתילציה של אתר ה-CpG ההמימתילציה יוצרת את אתר המחשוף של האנדונוקלאז Gla I – GCGC שעבר מתילציה מלאה. מחשוף Gla I של המוצר אוליגונוקלאוטיד משחרר את הפלואורופור מהקוונצ’ר ומייצר פלואורסצנטיות בזמן אמת. ניתן להשתמש בבדיקה כדי לבחון את הפעילות של כל איזופורם של DNMT; עם זאת, פעילות גבוהה יותר נצפית עם DNMT1 מכיוון שאיזוזים אלה מעדיפים מתילטים של דנ”אהמימתילציה 1,5. פעילות חזקה עוד יותר נצפתה אם התחום רצף מיקוד מוקדי שכפול אוטומטי (RFTS) מוסר מ- DNMT1. תחום זה, שנמצא באזור הרגולטורי N-terminal, נקשר לאתר הקטליטי ומונע קשירת DNA. הסרת ~600 חומצות האמינו הראשונות גורמת לאנזים חתוך שפעיל יותר באופן משמעותי מהאנזים באורך מלא (עלייה של ~פי 640 ב-kcat/Km)40. צורה פעילה זו של האנזים, המכונה DNMT1 חסר RFTS (חומצות אמינו 621-1616), מאפשרת זיהוי קל יותר של מעכבים בשל הכוח הקטליטי המוגבר שלו. מאמר זה מציג פרוטוקול לשימוש ב-DNMT1 חסר RFTS במבחנים כדי לסנן מעכבי מולקולות קטנות פוטנציאליות. באמצעות הבדיקה הרציפה המצומדת לאנדונוקלאז, המהירות הראשונית נקבעת בנוכחות ובהיעדר כמה מולקולות קטנות. כל מעכב פוטנציאלי נבדק בשני ריכוזים כדי לחפש עיכוב DNMT1 תלוי ריכוז. אחוז הפעילות שנצפתה בנוכחות המולקולות הקטנות חושב בכל מקרה.
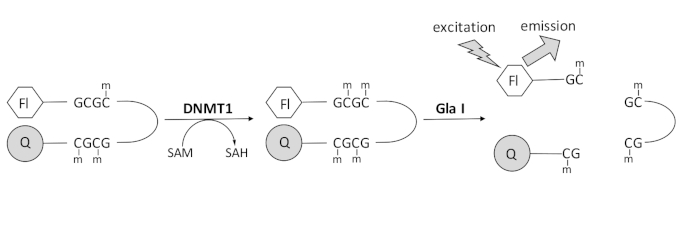
איור 1: בדיקת מתילציה של DNA. דנ”א של סיכות שיער המימתילציה עם פלואורופור בקצה 5′ וקוונצ’ר בקצה 3′ משמש כמצע. DNMT1 מזרז את העברת קבוצת המתיל מ-S-אדנוזיל-מתיונין לאתר CpG שאינו מתילציה, ויוצר S-אדנוזילהומוציסטאין ודנ“א שעבר מתילציה מלאה. מוצר הדנ”א מכיל את אתר הבקיעה של האנדונוקלאז Gla I, אשר מבקע אתרי GCGC שעברו מתילציה מלאה. ביקוע של דנ”א המוצר משחרר את הפלואורופור 5′ מהקוונצ’ר 3′, ויוצר פלואורסצנציה. קיצורים: Fl = פלואורופור; Q =quencher; DNMT1 = דנ”א מתיל-טרנספראז 1; SAM = S-אדנוזיל-מתיונין; SAH = S-אדנוזילהומוציסטאין. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
