A metilação do DNA é uma importante marca epigenética que regula a expressão gênica e a estrutura da cromatina. A metilação ocorre predominantemente nos dinucleotídeos CpG — citosina seguida de guanosina; o grupo metilo é adicionado à posição 5 da citosina. Padrões corretos de metilação do DNA e, portanto, a expressão gênica adequada, são necessários para o desenvolvimento e a função celular apropriados. Muitos estados patológicos têm sido associados a alterações no padrão normal de metilação 1,2,3. Por exemplo, existe uma ligação entre o início e a progressão do câncer e alterações no padrão de metilação do DNA. Normalmente, as células cancerosas exibem níveis gerais mais baixos de metilcitosina, o que contribui para a instabilidade do genoma. Ao mesmo tempo, a metilcitosina que está presente no genoma está concentrada nas regiões promotoras dos genes supressores de tumor, o que leva ao silenciamento genético dessas proteínas importantes. Notavelmente, as alterações epigenéticas são dinâmicas e reversíveis, ao contrário das mutações no DNA associadas à tumorigênese. Isso tornou as proteínas envolvidas na regulação epigenética gênica alvos de drogas interessantes 2,4.
As metiltransferases de DNA (DNMTs) são as proteínas responsáveis por gerar e manter os padrões de metilação do DNA. Três isozimas cataliticamente ativas, DNMT1, DNMT3a e DNMT3b, existem em humanos. Durante o desenvolvimento e diferenciação, as metiltransferases de novo, DNMT3a e DNMT3b, estabelecem padrões de metilação. Ambas as enzimas podem se ligar à proteína DNMT3L cataliticamente inativa para formar complexos que exibem atividade aumentada 1,5. Após a divisão celular, as células filhas contêm DNA hemimetilado – DNA contendo metilcitosina em apenas uma fita do duplex – porque o DNA recém-sintetizado é desprovido de marcas de metilação. A principal função do DNMT1 é metilar esse DNA hemimetilado, restabelecendo assim o padrão completo de metilação 1,5.
As ligações entre a atividade da DNMT e o câncer estão bem estabelecidas. A superexpressão de DNMT1, seja por mecanismos transcricionais ou pós-traducionais, é consequência de várias vias oncogênicas comuns 6,7,8,9. Abordagens genéticas para menor atividade da DNMT1 usando alelos hipomórficos resultam em diminuição da formação tumoral em camundongos Apc(Min)10. Os oligonucleotídeos antisense que derrubam a DNMT1 inibem a neoplasia em modelos de cultura celular e tumores de camundongos11,12. Assim, inibir a atividade da DNMT1 parece ser uma abordagem promissora de terapia contra o câncer. No entanto, os papéis que as isozimas DNMT3 desempenham não são tão simples. As mutações DNMT3a são encontradas na leucemia mieloide aguda13 e na síndrome mielodisplásica14. Pelo menos uma das mutações identificadas demonstrou diminuir a atividade de metilação do DNA da enzima15. No entanto, o DNMT3b é superexpresso no câncer de mama16 e no câncer colorretal17. Com as várias isozimas DNMT desempenhando papéis diferentes na carcinogênese, a identificação de inibidores específicos de isoenzimas será crítica. Não só esses compostos serão úteis para o desenvolvimento de terapias, mas os inibidores específicos da isoenzima também seriam uma ferramenta inestimável para dissecar o papel de cada isoenzima DNMT na etiologia do câncer.
Vários inibidores de DNMT têm sido relatados na literatura. Os inibidores conhecidos da DNMT podem ser divididos em duas classes: nucleosídeo e não nucleosídeo. Os inibidores de nucleosídeos são tipicamente análogos da citidina. Esses compostos são incorporados ao DNA e retêm covalentemente DNMTs. 5-azacitidina e 5-aza-2′-desoxicitidina foram aprovados para o tratamento da síndrome mielodisplásica e leucemia mieloide aguda 4,18. A alta toxicidade, a baixa biodisponibilidade e a instabilidade química desses compostos apresentam problemas. O trabalho em andamento está examinando a eficácia da próxima geração de inibidores de nucleosídeos; SGI-110, derivado de 5-aza-2′-desoxicitidina, é um exemplo19,20. Os inibidores de nucleosídeos não são específicos da isoenzima e inativarão qualquer isoenzima DNMT encontrada. Portanto, o tratamento com um agente nucleosídeo-desmetilante resulta na depleção de todas as isozimas DNMT 4,18. Os inibidores não nucleosídeos não precisam ser incorporados ao DNA para exercer seus efeitos inibitórios. Em vez disso, essas moléculas se ligam diretamente aos DNMTs, introduzindo a possibilidade de inibição específica da isozima. Vários inibidores não nucleosídeos foram descobertos até o momento, incluindo SGI-102721, hidralazina 22, procainamida 23, RG108 e derivados 24, e produtos naturais, (−)-epigalocatequina 3-galato (EGCG)25 e ácido lacáico A 26,27. A maioria dos inibidores não-nucleosídeos descobertos até o momento não são seletivos de isoenzima ou exibem preferências fracas para uma isoenzima DNMT. Além disso, a potência dessas moléculas precisa ser melhorada, principalmente nas células 4,18. Assim, há uma necessidade de descobrir ou desenvolver inibidores de DNMT mais potentes e seletivos de isozima.
Um obstáculo para a descoberta de novos inibidores de pequenas moléculas de DNMTs são os ensaios laboriosos tradicionalmente usados para examinar a atividade da DNMT28. Os ensaios geralmente são descontínuos com várias etapas. A atividade enzimática dos DNMTs ainda é rotineiramente ensaiada utilizando S-adenosilmetionina radioativa (SAM)29,30,31,32,33,34. Ensaios não radioativos para metilação do DNA também foram desenvolvidos. Por exemplo, ensaios utilizando endonucleases de restrição sensíveis à metila e eletroforese para separar os produtos da digestão têm sido descritos35,36. Esses tipos de ensaios descontínuos e em várias etapas não são facilmente passíveis de descoberta de medicamentos. Desde meados dos anos 2000, vários ensaios de metilação de DNA com maior rendimento foram desenvolvidos28. Um ensaio de proximidade de cintilação foi utilizado para triagem de inibidores de DNMT137. Outro ensaio utilizando endonuclease de restrição sensível à metila foi utilizado para rastrear inibidores de DNMT3a25,38. Embora ambos os ensaios tenham permitido um rendimento mais elevado do que os ensaios tradicionais de metilação do ADN, os ensaios requerem várias etapas e não permitem a observação da atividade de metilação em tempo real. Mais recentemente, um ensaio de cinética contínua tem sido descrito que acopla a formação de S-adenosilhomocisteína (HAS), um produto da reação de metilação, à alteração espectroscópica a 340 nm associada à oxidação do NADPH39. Este ensaio utiliza três enzimas de acoplamento para gerar um sinal espectroscópico.
Desenvolvemos um ensaio de metilação de DNA acoplado à endonuclease baseado em fluorescência que utiliza uma única enzima de acoplamento comercialmente disponível e pode gerar dados em tempo real (Figura 1). Um oligonucleotídeo hairpin contendo três metilcitosinas é usado como substrato. O DNA do substrato contém um fluoróforo na extremidade 5′ e um quencher na extremidade 3′. A metilação do sítio CpG hemimetilado gera o sítio de clivagem para a endonuclease Gla I — GCGC totalmente metilado. A clivagem Gla I do oligonucleotídeo do produto libera o fluoróforo do quencher e gera fluorescência em tempo real. O ensaio pode ser usado para examinar a atividade de qualquer isoforma de DNMT; no entanto, observa-se maior atividade com DNMT1, pois esta isoenzima preferencialmente metila o DNA hemimetilado 1,5. Atividade ainda mais robusta é observada se o domínio RFTS (Replication Foci Targeting Sequence) autoinibitório for removido da DNMT1. Este domínio, encontrado na região reguladora N-terminal, liga-se ao sítio catalítico e impede a ligação do DNA. A remoção dos primeiros ~600 aminoácidos resulta em uma enzima truncada que é significativamente mais ativa do que a enzima de comprimento total (~640 vezes maior em kcat/Km)40. Esta forma ativada da enzima, referida como DNMT1 sem RFTS (aminoácidos 621-1616), permite a identificação mais fácil de inibidores devido ao seu aumento do poder catalítico. Este artigo apresenta um protocolo para utilizar DNMT1 sem RFTS em ensaios para triagem de potenciais inibidores de moléculas pequenas. Usando o ensaio contínuo acoplado à endonuclease, a velocidade inicial é determinada na presença e ausência de algumas moléculas pequenas. Cada inibidor potencial é examinado em duas concentrações para procurar inibição DNMT1 dependente da concentração. A porcentagem de atividade observada na presença das pequenas moléculas foi calculada em cada caso.
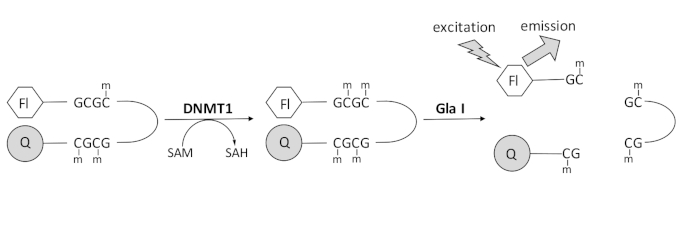
Figura 1: Ensaio de metilação do DNA. Um DNA de grampo de cabelo hemimetilado com um fluoróforo na extremidade 5′ e um quencher na extremidade 3′ é usado como substrato. DNMT1 catalisa a transferência do grupo metilo da S-adenosilmetionina para o sítio CpG não metilado, gerando S-adenosilhomocisteína e DNA totalmente metilado. O produto de DNA contém o local de clivagem para a endonuclease Gla I, que cliva os sítios GCGC totalmente metilados. A clivagem do DNA do produto libera o fluoróforo de 5′ do quencher de 3′, gerando fluorescência. Abreviaturas: Fl = fluoróforo; Q = quencher; DNMT1 = DNA metiltransferase 1; SAM = S-adenosilmetionina; HAS = S-adenosilhomocisteína. Por favor, clique aqui para ver uma versão maior desta figura.
