Метилирование ДНК является важным эпигенетическим маркером, который регулирует экспрессию генов и структуру хроматина. Метилирование происходит преимущественно в cpG динуклеотидах — цитозине, за которым следует гуанозин; метильную группу добавляют к 5-позиции цитозина. Правильные паттерны метилирования ДНК и, следовательно, правильная экспрессия генов необходимы для соответствующего клеточного развития и функционирования. Многие болезненные состояния были связаны с изменениями нормальной картины метилирования 1,2,3. Например, существует связь между инициацией и прогрессированием рака и изменениями в паттерне метилирования ДНК. Как правило, раковые клетки демонстрируют более низкие общие уровни метилцитозина, что способствует нестабильности генома. При этом метилцитозин, присутствующий в геноме, концентрируется в промоторных областях генов-супрессоров опухолей, что приводит к глушению генов этих важных белков. Примечательно, что эпигенетические изменения являются динамичными и обратимыми, в отличие от мутаций ДНК, связанных с опухолевым генезом. Это сделало белки, участвующие в эпигенетической регуляции генов, интересными лекарственными мишенями 2,4.
ДНК-метилтрансферазы (DDNMT) являются белками, ответственными за генерацию и поддержание паттернов метилирования ДНК. Три каталитически активных изофермента, DNMT1, DNMT3a и DNMT3b, существуют у людей. Во время разработки и дифференцировки de novo метилтрансферазы, DNMT3a и DNMT3b, устанавливают паттерны метилирования. Оба фермента могут связывать каталитически неактивный белок DNMT3L с образованием комплексов, которые проявляют повышенную активность 1,5. После деления клеток дочерние клетки содержат гемиметилированную ДНК — ДНК, содержащую метилцитозин только в одной нити дуплекса — потому что вновь синтезированная ДНК лишена меток метилирования. Основная функция DNMT1 заключается в метилировании этой гемиметилированной ДНК, тем самым восстанавливая полную картину метилирования 1,5.
Связи между активностью DNMT и раком хорошо известны. Сверхэкспрессия DNMT1, либо транскрипционными, либо посттрансляционными механизмами, является следствием нескольких общих онкогенных путей 6,7,8,9. Генетические подходы к снижению активности DNMT1 с использованием гипоморфных аллелей приводят к уменьшению образования опухоли у мышей Apc(Min)10. Антисмысловые олигонуклеотиды, которые нокдают DNMT1, ингибируют неоплазию в клеточной культуре и моделях опухолей мышей 11,12. Таким образом, ингибирование активности DNMT1 кажется многообещающим подходом к терапии рака. Однако роли, которые играют изоферменты DNMT3, не так просты. Мутации DNMT3a обнаруживаются при остром миелоидном лейкозе13 и миелодиспластическом синдроме14. Было показано, что по меньшей мере одна из идентифицированных мутаций снижает активность метилирования ДНК фермента15. Тем не менее, DNMT3b чрезмерно экспрессируется при раке молочной железы16 и колоректальном раке17. Поскольку различные изоферменты DNMT играют различную роль в канцерогенезе, идентификация изофермент-специфических ингибиторов будет иметь решающее значение. Мало того, что эти соединения будут полезны для разработки терапевтических средств, но изофермент-специфические ингибиторы также будут бесценным инструментом для анализа роли каждого изофермента DNMT в этиологии рака.
В литературе сообщалось о нескольких ингибиторах DNMT. Известные ингибиторы DNMT можно разделить на два класса: нуклеозидные и ненуклеозидные. Нуклеозидные ингибиторы обычно являются аналогами цитидина. Эти соединения включены в ДНК и ковалентно улавливают DNMTs. 5-азацитидин и 5-аза-2′-дезоксицитидин были одобрены для лечения миелодиспластического синдрома и острого миелоидного лейкоза 4,18. Высокая токсичность, низкая биодоступность и химическая нестабильность этих соединений представляют проблемы. Текущая работа изучает эффективность следующего поколения нуклеозидных ингибиторов; SGI-110, полученный из 5-аза-2′-дезоксицитидина, является одним из примеров19,20. Нуклеозидные ингибиторы не являются изофермент-специфическими и инактивируют любой встречающийся изофермент DNMT. Таким образом, обработка нуклеозид-деметилирующим агентом приводит к истощению всех изоферментов DNMT 4,18. Ненуклеозидные ингибиторы не нужно включать в ДНК, чтобы оказывать их ингибирующее действие. Вместо этого эти молекулы связываются непосредственно с DDNMT, вводя возможность для изофермент-специфического ингибирования. На сегодняшний день было обнаружено несколько ненуклеозидных ингибиторов, включая SGI-102721, гидралазин22, прокаинамид23, RG108 и производные24, а также натуральные продукты, (-)-эпигаллокатехин 3-галлат (EGCG)25 и лаккаевую кислоту A26,27. Большинство ненуклеозидных ингибиторов, обнаруженных на сегодняшний день, не являются изофермент-селективными или демонстрируют слабые предпочтения для одного изофермента DNMT. Кроме того, необходимо улучшить эффективность этих молекул, особенно в клетках 4,18. Таким образом, существует необходимость в открытии или разработке более мощных, изофермент-селективных ингибиторов DNMT.
Препятствием для открытия новых низкомолекулярных ингибиторов DNMT являются трудоемкие анализы, традиционно используемые для изучения активности DNMT28. Анализы обычно прерывистые с несколькими шагами. Ферментативная активность DNMT по-прежнему регулярно анализируется с использованием радиоактивного S-аденозилметионина (SAM)29,30,31,32,33,34. Также были разработаны нерадиоактивные анализы метилирования ДНК. Например, описаны анализы с использованием метилчувствительных эндонуклеаз рестрикции и электрофореза для разделения продуктов пищеварения35,36. Эти типы прерывистых, многоступенчатых анализов не поддаются открытию лекарств. С середины 2000-х годов было разработано несколько анализов метилирования ДНК с более высокой пропускной способностью28. Сцинтилляционный бесконтактный анализ использовался для скрининга ингибиторов DNMT137. Другой анализ с использованием метил-чувствительной рестрикционной эндонуклеазы был использован для скрининга ингибиторов DNMT3a25,38. Хотя оба анализа позволили получить более высокую пропускную способность, чем традиционные анализы метилирования ДНК, анализы требуют нескольких этапов и не позволяют наблюдать активность метилирования в режиме реального времени. Совсем недавно был описан непрерывный кинетический анализ, который связывает образование S-аденозилгомоцистеина (SAH), одного из продуктов реакции метилирования, со спектроскопическим изменением при 340 нм, связанным с окислением NADPH39. Этот анализ использует три фермента связи для генерации спектроскопического сигнала.
Мы разработали анализ метилирования ДНК на основе флуоресценции, связанный с эндонуклеазой, который использует один коммерчески доступный фермент связи и может генерировать данные в режиме реального времени (рисунок 1). В качестве субстрата используется олигонуклеотид шпильки, содержащий три метилцитозина. СУБСТРАТНАЯ ДНК содержит флуорофор на 5′ конце и гаситель на 3′ конце. Метилирование гемиметилированного сайта CpG генерирует участок расщепления для эндонуклеазы Gla I — полностью метилированного GCGC. Расщепление продукта олигонуклеотид gla I высвобождает флуорофор из гасителя и генерирует флуоресценцию в режиме реального времени. Анализ может быть использован для изучения активности любой изоформы DNMT; однако более высокая активность наблюдается при DNMT1, так как этот изофермент преимущественно метилирует гемиметилированную ДНК 1,5. Еще более устойчивая активность наблюдается, если домен автоингибаторной последовательности таргетирования фокусов репликации (RFTS) удаляется из DNMT1. Этот домен, обнаруженный в N-концевой регуляторной области, связывается с каталитическим сайтом и предотвращает связывание ДНК. Удаление первых ~ 600 аминокислот приводит к усеченному ферменту, который значительно более активен, чем фермент полной длины (~ 640-кратное увеличение kcat/Km)40. Эта активированная форма фермента, называемая RFTS-отсутствующим DNMT1 (аминокислоты 621–1616), позволяет легче идентифицировать ингибиторы из-за его повышенной каталитической мощности. В этой статье представлен протокол для использования RFTS-отсутствующего DNMT1 в анализах для скрининга потенциальных ингибиторов малых молекул. С помощью непрерывного анализа, связанного с эндонуклеазой, начальную скорость определяют в присутствии и отсутствии нескольких малых молекул. Каждый потенциальный ингибитор исследуется в двух концентрациях для поиска концентрационно-зависимого ингибирования DNMT1. Процент активности, наблюдаемый в присутствии малых молекул, был рассчитан в каждом случае.
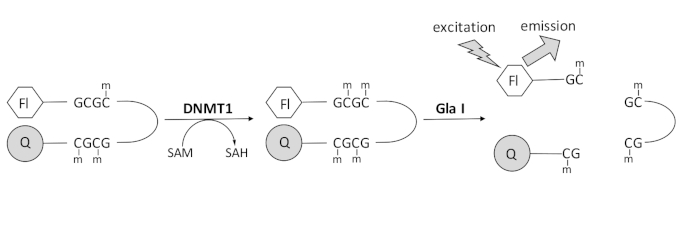
Рисунок 1: Анализ метилирования ДНК. В качестве субстрата используется гемиметилированная шпилька ДНК с флуорофором на 5′ конце и гасителем на 3′ конце. DNMT1 катализирует перенос метильной группы из S-аденозилметионина в неметилированный сайт CpG, генерируя S-аденозилгомоцистеин и полностью метилированную ДНК. Продукт ДНК содержит участок расщепления эндонуклеазы Gla I, которая расщепляет полностью метилированные участки GCGC. Расщепление ДНК продукта высвобождает 5′ флуорофор из 3′ гасителя, генерируя флуоресценцию. Сокращения: Fl = флуорофор; Q = quencher; DNMT1 = ДНК-метилтрансфераза 1; SAM = S-аденозилметионин; SAH = S-аденозилгомоцистеин. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
