L’optogénétique, le contrôle des processus biologiques avec des protéines sensibles à la lumière, offre une nouvelle stratégie pour contrôler dynamiquement les fermentations microbiennes pour la production chimique et protéique1,2. Le fardeau des voies métaboliques modifiées et la toxicité de certains intermédiaires et produits nuisent souvent à la croissance cellulaire3. De tels stress peuvent entraîner une mauvaise accumulation de biomasse et une productivité réduite3. Ce défi peut être relevé en divisant temporellement les fermentations en une phase de croissance et de production, qui consacrent des ressources métaboliques à l’accumulation de biomasse ou à la synthèse de produits respectivement4. Nous avons récemment montré que la transition de la croissance à la production dans cette fermentation en deux phases peut être induite par des changements dans les conditions d’éclairage5,6,7. La haute accordabilité, la réversibilité et l’orthogonalité des apports lumineux8 offrent des avantages uniques aux fermentations contrôlées par la lumière qui sont difficiles ou impossibles à reproduire avec des inducteurs chimiques utilisés dans le contrôle dynamique des fermentations biphasées conventionnelles4,9,10,11.
La protéine EL222 sensible à la lumière bleue dérivée d’Erythrobacter litoralis a été utilisée pour développer plusieurs circuits optogénétiques pour l’ingénierie métabolique chez Saccharomyces cerevisiae5,7,12,13. EL222 contient un domaine de capteur lumière-oxygène-tension (LOV) qui subit un décalage conformationnel lors de l’activation de la lumière bleue (465 nm), ce qui lui permet de se lier à sa séquence d’ADN apparenté (C120)13. La fusion d’EL222 au domaine d’activation viral VP16 (VP16-EL222) donne un facteur de transcription sensible à la lumière bleue qui peut activer de manière réversible l’expression génique chez S. cerevisiae7 et d’autres organismes14 à partir du promoteur synthétique PC120. Plusieurs circuits basés sur EL222 ont été développés et utilisés pour la production chimique chez S. cerevisiae, tels que le système OptoEXP de base activé par la lumière7, dans lequel le gène d’intérêt est directement exprimé à partir de PC120 (Figure 1A). Cependant, les préoccupations de pénétration de la lumière aux densités cellulaires élevées généralement rencontrées dans la phase de production des fermentations nous ont motivés à développer des circuits inversés induits dans l’obscurité, tels que les circuits OptoINVRT et OptoQ-INVRT (Figure 1B)5,7,13. Ces systèmes exploitent les régulons de galactose (GAL) ou d’acide quinique (Q) de S. cerevisiae et N. crassa, respectivement, contrôlant leurs répresseurs correspondants (GAL80 et QS) avec VP16-EL222, pour réprimer l’expression des gènes dans la lumière et l’induire fortement dans l’obscurité. La combinaison des circuits OptoEXP et OptoINVRT permet un contrôle bidirectionnel de l’expression des gènes, permettant des fermentations en deux phases dans lesquelles la phase de croissance est induite par la lumière bleue et la phase de production par l’obscurité (Figure 2A)5,7.
L’utilisation de la lumière au lieu de l’obscurité pour induire l’expression des gènes pendant la phase de production élargirait considérablement les capacités des contrôles optogénétiques, mais nécessiterait également de surmonter les limites de pénétration de la lumière des densités cellulaires élevées généralement rencontrées dans cette phase de fermentation. À cette fin, nous avons développé des circuits, connus sous le nom d’OptoAMP et OptoQ-AMP, qui amplifient la réponse transcriptionnelle à la stimulation par la lumière bleue. Ces circuits utilisent des mutants de type sauvage ou hypersensibles de VP16-EL222 pour contrôler la production des activateurs transcriptionnels Gal4p ou QF2 des régulons GAL ou Q, respectivement, obtenant une sensibilité accrue et une expression génique plus forte avec la lumière12,13 (Figure 1C). Les circuits OptoAMP peuvent obtenir une induction lumineuse complète et homogène dans des bioréacteurs de 5 L à une densité optique (mesurée à 600 nm; OD600) valeurs d’au moins 40 avec seulement ~ 0,35% d’éclairage (dose de lumière de 5% sur seulement ~ 7% de la surface en vrac). Cela démontre un degré de sensibilité plus élevé par rapport à OptoEXP, qui nécessite un éclairage proche de 100 %12. La capacité d’induire efficacement l’expression des gènes avec de la lumière à des densités cellulaires élevées ouvre de nouvelles possibilités de contrôle dynamique des fermentations. Cela inclut l’exploitation de fermentations dans plus de deux phases temporelles, telles que les fermentations triphasées, dans lesquelles les phases de croissance, d’induction et de production sont établies avec des calendriers d’éclairage uniques pour optimiser la production chimique (Figure 2B)12.
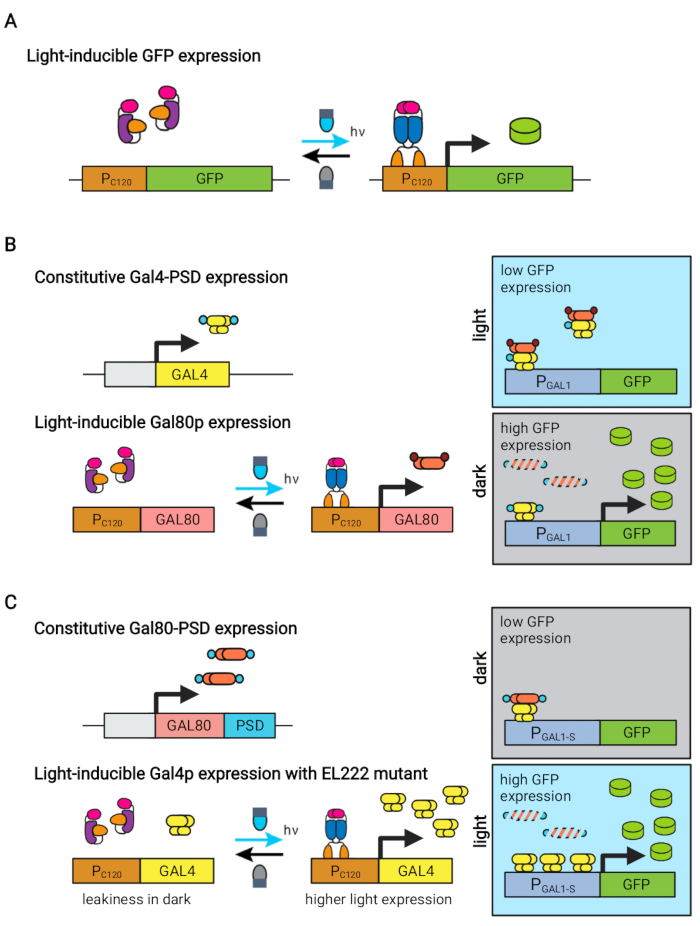
Figure 1 : Circuits optogénétiques pour le contrôle dynamique de S. cerevisiae. Les circuits OptoEXP, OptoINVRT et OptoAMP sont basés sur le système sensible à la lumière VP16-EL222. (A) Dans le circuit OptoEXP, l’exposition à la lumière bleue provoque un changement conformationnel et une dimérisation de VP16-EL222, ce qui expose un domaine de liaison à l’ADN et permet la transcription à partir de PC120. La figure a été modifiée à partir de Zhao et al.7. (B) Les circuits OptoINVRT exploitent les régulons GAL (illustrés) ou Q pour induire une expression dans l’obscurité. Dans les circuits à base de GAL, VP16-EL222 et GAL4 sont exprimés de manière constitutive, tandis que le PC120 entraîne l’expression du répresseur GAL80 (dans les circuits à base de Q, GAL4 et GAL80 sont remplacés par QF2 et QS, respectivement, et un promoteur synthétique contenant DU QUAS est utilisé à la place d’un promoteur GAL). À la lumière, Gal80p empêche l’activation du gène d’intérêt de PGAL1. Dans l’obscurité, GAL80 n’est pas exprimé et se dégrade rapidement en le fusionnant en un domaine degron constitutif (petit domaine brun), ce qui permet l’activation de PGAL1 par Gal4p. La figure a été modifiée à partir de Zhao et al.5. (C) Les circuits OptoAMP utilisent également VP16-EL222 pour contrôler les régulons GAL (illustrés) ou Q. Dans ces circuits, le répresseur GAL80 (ou QS) est exprimé de manière constitutive et fusionné à un degron photosensible (petit domaine bleu) assurant une répression stricte dans l’obscurité. PC120 et un mutant hypersensible VP16-EL222 contrôlent l’expression de GAL4 (ou QF2) avec la lumière, ce qui active fortement PGAL1 (ou un promoteur contenant QUAS) dans la lumière. Les circuits dérivés de GAL peuvent utiliser des formes techniques de PGAL1, telles que PGAL1-M ou PGAL1-S, qui ont une activité accrue, ainsi que des promoteurs de type sauvage contrôlés par le régulateur GAL (PGAL1, PGAL10, PGAL2, PGAL7). La figure a été modifiée à partir de Zhao et al.12. Veuillez cliquer ici pour voir une version plus grande de cette figure.
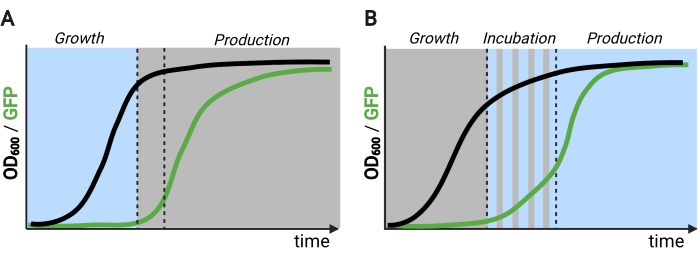
Figure 2: Fermentations biphasées et triphasées dans le temps. (A) Les fermentations biphasées opérées avec des circuits inversés se composent d’une phase de croissance entraînée par la lumière et d’une phase de production sombre. Dans la phase de croissance, la biomasse s’accumule à mesure que la voie de production reste réprimée. Lorsqu’elles atteignent l’OD600 souhaitée, les cellules sont déplacées vers l’obscurité pour s’ajuster métaboliquement avant d’être remises en suspension dans des milieux frais pour la phase de production. (B) Dans un processus en trois phases, les phases de croissance, d’incubation et de production sont définies par des horaires lumineux uniques, qui peuvent consister en une période de croissance sombre, une incubation pulsée et une phase de production entièrement éclairée. Figure créée avec Biorender. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Des circuits optogénétiques ont également été développés pour le contrôle dynamique de la production chimique et protéique chez E. coli. Les circuits OptoLAC contrôlent le répresseur LacI bactérien à l’aide du circuit pDawn sensible à la lumière, basé sur le système à deux composants YF1/FixJ6 (Figure 3). Semblables à OptoINVRT5, les circuits OptoLAC sont conçus pour réprimer l’expression des gènes dans la lumière et l’induire dans l’obscurité. Les niveaux d’expression utilisant les circuits OptoLAC peuvent égaler ou dépasser ceux atteints avec l’induction standard de l’isopropyle β-d-1-thiogalactopyranoside (IPTG), maintenant ainsi la force de l’induction chimique tout en offrant une accordabilité et une réversibilité améliorées6. Par conséquent, les circuits OptoLAC permettent un contrôle optogénétique efficace pour l’ingénierie métabolique chez E. coli.
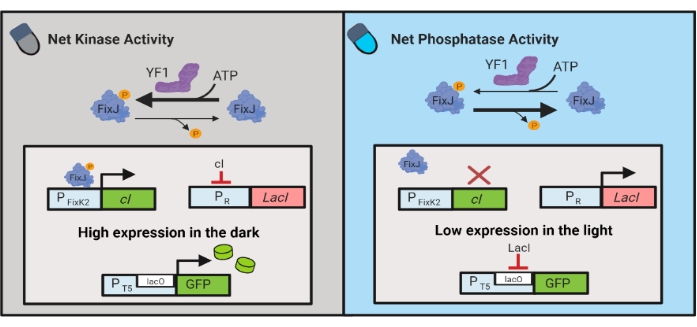
Figure 3 : Circuits OptoLAC pour le contrôle dynamique d’E. coli. Les circuits OptoLAC adaptent le système pDawn et l’opéron lac pour réaliser l’activation dans l’obscurité et la répression dans la lumière. Dans l’obscurité, YF1 phosphoryle FixJ, qui active ensuite le promoteur PFixK2 pour exprimer le répresseur cI. Le répresseur cI empêche l’expression du répresseur lacI à partir du promoteur PR, ce qui permet la transcription du gène d’intérêt à partir d’un promoteur contenant du lacO. Inversement, la lumière bleue réduit l’activité de la kinase nette YF1, inversant la phosphorylation fixJ et donc l’expression cI, ce qui déprime l’expression de lacI et empêche l’expression du promoteur contenant du lacO. La figure a été modifiée à partir de Lalwani et al.6. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Nous décrivons ici les protocoles de base pour les fermentations contrôlées par la lumière de S. cerevisiae et E. coli pour la production chimique ou protéique. Pour les levures et les bactéries, nous nous concentrons d’abord sur les fermentations avec une phase de croissance entraînée par la lumière et une phase de production induite par l’obscurité rendue possible par les circuits OptoINVRT et OptoLAC. Par la suite, nous décrivons un protocole pour une fermentation triphasée (croissance, induction, production) contrôlée par la lumière rendue possible par les circuits OptoAMP. En outre, nous décrivons comment étendre les fermentations contrôlées optogénétiquement des microplaques aux bioréacteurs à l’échelle du laboratoire. Avec ce protocole, nous visons à fournir un guide complet et facilement reproductible pour effectuer des fermentations contrôlées par la lumière pour la production de produits chimiques ou de protéines.
