Оптогенетика, контроль биологических процессов с помощью светочувствительных белков, предлагает новую стратегию динамического контроля микробных ферментаций для химического и белкового производства1,2. Бремя инженерных метаболических путей и токсичность некоторых промежуточных продуктов и продуктов часто ухудшают рост клеток3. Такие стрессы могут привести к плохому накоплению биомассы и снижению производительности3. Эта проблема может быть решена путем временного разделения ферментаций на фазу роста и производства, которые выделяют метаболические ресурсы для накопления биомассы или синтеза продукта соответственно4. Недавно мы показали, что переход от роста к производству в этой двухфазной ферментации может быть вызван изменением условий освещения5,6,7. Высокая настраиваемость, обратимость и ортогональность световых входов8 обеспечивают уникальные преимущества для контролируемых светом ферментаций, которые трудно или невозможно воспроизвести с помощью химических индукторов, используемых в динамическом управлении обычными двухфазными ферментациями4,9,10,11.
Чувствительный к синему свету белок EL222, полученный из Erythrobacter litoralis, был использован для разработки нескольких оптогенетических схем для метаболической инженерии в Saccharomyces cerevisiae5,7,12,13. EL222 содержит домен датчика напряжения света и кислорода (LOV), который претерпевает конформационный сдвиг при активации синего света (465 нм), что позволяет ему связываться с его родственной последовательностью ДНК (C120)13. Слияние EL222 с вирусным доменом активации VP16 (VP16-EL222) приводит к созданию фактора транскрипции, реагирующего на синий свет, который может обратимо активировать экспрессию генов у S. cerevisiae7 и других организмов14 из синтетического промотора PC120. Несколько схем на основе EL222 были разработаны и использованы для химического производства в S. cerevisiae, такие как базовая светоактивированная система OptoEXP7, в которой интересующий ген непосредственно экспрессируется из PC120 (рисунок 1A). Тем не менее, опасения по поводу проникновения света при высокой плотности клеток, обычно встречающиеся на производственной фазе ферментаций, побудили нас разработать перевернутые цепи, которые индуцируются в темноте, такие как схемы OptoINVRT и OptoQ-INVRT (рисунок 1B) 5,7,13. Эти системы используют регулоны галактозы (GAL) или хиновой кислоты (Q) из S. cerevisiae и N. crassa соответственно, контролируя их соответствующие репрессоры (GAL80 и QS) с помощью VP16-EL222, чтобы подавить экспрессию генов на свету и сильно индуцировать ее в темноте. Объединение схем OptoEXP и OptoINVRT приводит к двунаправленному контролю экспрессии генов, что позволяет проводить двухфазные ферментации, в которых фаза роста индуцируется синим светом, а фаза производства — темнотой (рисунок 2A)5,7.
Использование света вместо темноты для индуцирования экспрессии генов во время фазы производства значительно расширит возможности оптогенетического контроля, но также потребует преодоления ограничений проникновения света из-за высокой плотности клеток, обычно встречающихся в этой фазе ферментации. С этой целью мы разработали схемы, известные как OptoAMP и OptoQ-AMP, которые усиливают транскрипционный ответ на стимуляцию синего света. Эти схемы используют дикие или гиперчувствительные мутанты VP16-EL222 для контроля производства транскрипционных активаторов Gal4p или QF2 регулонов GAL или Q соответственно, достигая повышенной чувствительности и более сильной экспрессии генов с помощью light12,13 (рисунок 1C). Контуры OptoAMP могут достигать полной и однородной световой индукции в 5 л биореакторах при оптической плотности (измеренной при 600 нм; OD600) значения не менее 40 при освещении всего ~0,35% (5% дозы света только на ~7% объемной поверхности). Это демонстрирует более высокую степень чувствительности по сравнению с OptoEXP, который требует почти 100% освещения12. Способность эффективно индуцировать экспрессию генов светом при высокой плотности клеток открывает новые возможности для динамического контроля ферментаций. Это включает в себя рабочие ферментации в более чем двух временных фазах, таких как трехфазные ферментации, в которых фазы роста, индукции и производства устанавливаются с уникальными световыми графиками для оптимизации химического производства (рисунок 2B)12.
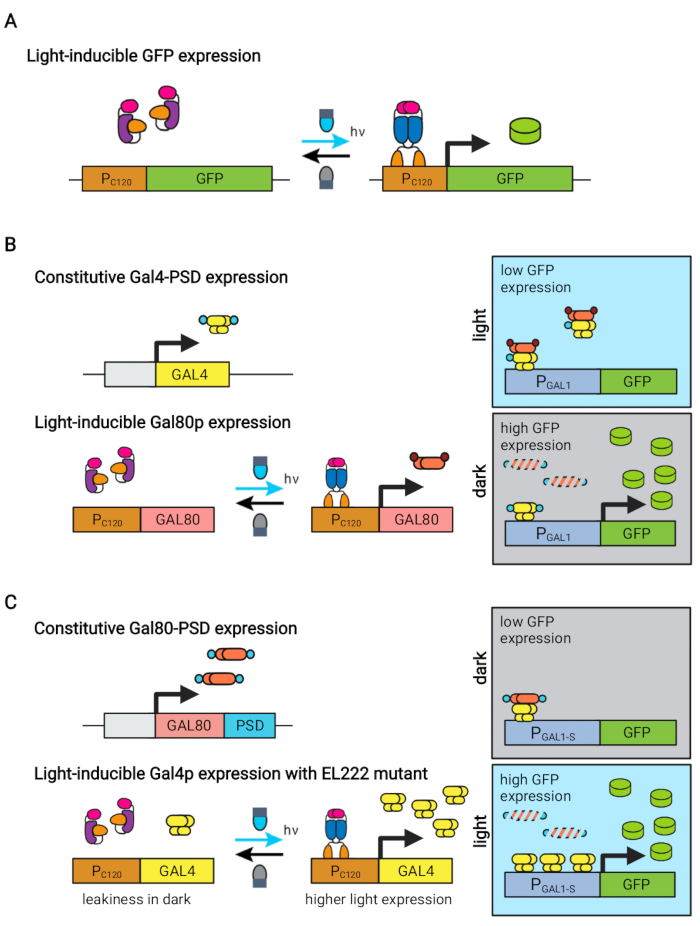
Рисунок 1: Оптогенетические схемы динамического управления S. cerevisiae. Схемы OptoEXP, OptoINVRT и OptoAMP основаны на светочувствительной системе VP16-EL222. (A) В цепи OptoEXP воздействие синего света вызывает конформационное изменение и димеризацию VP16-EL222, что обнажает ДНК-связывающий домен и позволяет транскрипцию из PC120. Рисунок был изменен по сравнению с Zhao et al.7. (B) Контуры OptoINVRT используют GAL (показаны) или Q регулоны для индуцирования экспрессии в темноте. В схемах на основе GAL ПОСЛЕДОВАТЕЛЬНО выражены VP16-EL222 и GAL4 , в то время как PC120 управляет экспрессией репрессора GAL80 (в схемах на основе Q GAL4 и GAL80 заменяются QF2 и QS соответственно, а вместо промотора GAL используется синтетический ПРОМОТОР, содержащий QUAS). В свете Gal80p предотвращает активацию интересующего гена из PGAL1. В темноте GAL80 не выражается и быстро деградирует путем слияния его с конститутивным дегронным доменом (малый коричневый домен), что позволяет активировать PGAL1 Gal4p. Рисунок был изменен по сравнению с Чжао и др.5. (C) Схемы OptoAMP также используют VP16-EL222 для управления GAL (показаны) или Q-регулонами. В этих схемах репрессор GAL80 (или QS) конститутивно выражен и слит с фоточувствительным дегроном (малый синий домен), обеспечивающим жесткое подавление в темноте. PC120 и гиперчувствительная VP16-EL222 мутантная контрольная экспрессия GAL4 (или QF2) со светом, который сильно активирует PGAL1 (или QUAS-содержащий промотор) в свете. Схемы, полученные из GAL, могут использовать инженерные формы PGAL1, такие как PGAL1-M или PGAL1-S, которые имеют повышенную активность, а также промоторы дикого типа, контролируемые регулятором GAL (PGAL1, PGAL10, PGAL2, PGAL7). Рисунок был изменен из Zhao et al.12. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
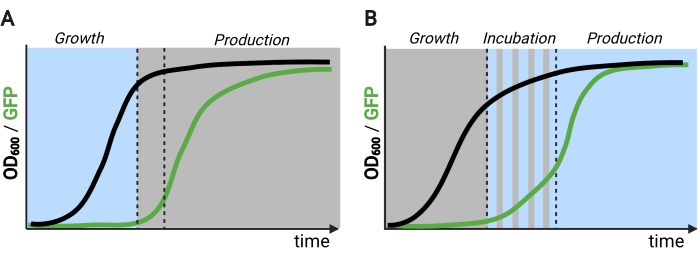
Рисунок 2: Двух- и трехфазные ферментации во времени. (А) Двухфазные ферментации, работающие с перевернутыми контурами, состоят из фазы роста, управляемой светом, и темной фазы производства. В фазе роста биомасса накапливается по мере того, как производственный путь остается подавленным. По достижении желаемого OD600 клетки смещаются в темноту для метаболической корректировки, прежде чем быть повторно суспендированными в свежих средах для фазы производства. (B) В трехфазном процессе фазы роста, инкубации и производства определяются уникальными световыми графиками, которые могут состоять из темного периода роста, импульсной инкубации и полностью освещенной фазы производства. Рисунок, созданный с помощью Biorender. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Также были разработаны оптогенетические схемы для динамического контроля химического и белкового производства в кишечной палочке. Схемы OptoLAC управляют бактериальным репрессором LacI с помощью светочувствительной схемы pDawn, которая основана на двухкомпонентной системе YF1/FixJ6 (рисунок 3). Подобно OptoINVRT5, схемы OptoLAC предназначены для подавления экспрессии генов на свету и индуцирования ее в темноте. Уровни экспрессии с использованием схем OptoLAC могут совпадать или превышать уровни, достигнутые при стандартной индукции изопропил-β-d-1-тиогалактопиранозида (IPTG), тем самым сохраняя силу химической индукции, обеспечивая при этом повышенную настраиваемость и обратимость6. Таким образом, схемы OptoLAC обеспечивают эффективный оптогенетический контроль метаболической инженерии в E. coli.
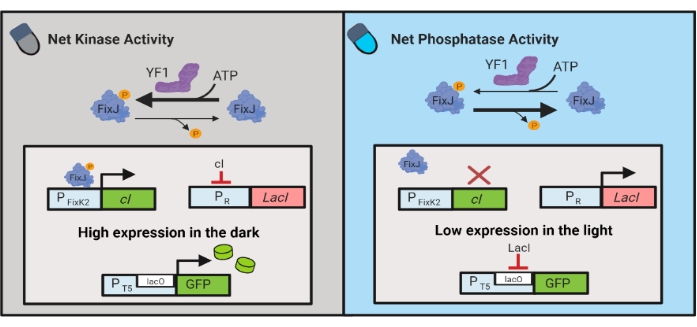
Рисунок 3: Схемы OptoLAC для динамического управления кишечной палочкой. Схемы OptoLAC адаптируют систему pDawn и lac operon для достижения активации в темноте и подавления на свету. В темноте YF1 фосфорилирует FixJ, который затем активирует промотор PFixK2 для экспрессии cI-репрессора . CI-репрессор предотвращает экспрессию репрессора lacI от промотора PR , что позволяет транскрипцию интересующего гена от lacO-содержащего промотора. И наоборот, синий свет снижает активность чистой киназы YF1, обращая вспять фосфорилирование FixJ и, следовательно, экспрессию cI , что снижает экспрессию lacI и предотвращает экспрессию от lacO-содержащего промотора. Эта цифра была изменена по сравнению с Лалвани и др.6. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Здесь мы опишем основные протоколы для контролируемых светом ферментаций S. cerevisiae и E. coli для химического или белкового производства. Как для дрожжей, так и для бактерий мы сначала фокусируемся на ферментациях со световой фазой роста и фазой производства, вызванной темнотой, обеспечиваемой схемами OptoINVRT и OptoLAC. Далее мы описываем протокол для трехфазной (рост, индукция, производство) легко контролируемой ферментации, обеспечиваемой схемами OptoAMP. Кроме того, мы описываем, как масштабировать оптогенетически контролируемые ферментации от микропластин до лабораторных биореакторов. С помощью этого протокола мы стремимся предоставить полное и легко воспроизводимое руководство для выполнения контролируемых светом ферментаций для химического или белкового производства.
