מטרת שיטה זו היא לחקור כיצד קלט רשתית משפיע על הגדילה וההתפתחות של טקטום אופטי, מרכז העיבוד החזותי במוח דגי הזברה. על ידי הסרת עין אחת ולאחר מכן השוואת שני הצדדים של טקטום אופטי, ניתן לצפות ולנרמל שינויים טקטליים בתוך אותה דגימה, מה שמאפשר השוואה בין דגימות מרובות. גישות מולקולריות מודרניות בשילוב עם טכניקה זו יניבו תובנות על המנגנונים העומדים בבסיס הצמיחה וההתפתחות של מערכת הראייה, כמו גם על ניוון והתחדשות אקסונאלית.
מערכות חושיות – חזותיות, שמיעתיות וסומטוסנסוריות – אוספות מידע מאיברים חיצוניים ומעבירות את המידע הזה למערכת העצבים המרכזית, ויוצרות “מפות” של העולם החיצוני על פני המוח האמצעי 1,2. ראייה היא השיטה החושית הדומיננטית עבור כמעט כל בעלי החוליות, כולל דגים רבים. הרשתית, הרקמה העצבית בעין, אוספת מידע עם מעגל עצבי המורכב בעיקר מפוטורצפטורים, תאים דו קוטביים ותאי גנגליון רשתית (RGCs), נוירוני ההקרנה של הרשתית. ל-RGCs יש אקסונים ארוכים שמוצאים את דרכם על פני השטח הפנימיים של הרשתית אל ראש עצב הראייה, שם הם מתפתלים ונעים יחד דרך המוח, ובסופו של דבר מסתיימים במרכז העיבוד החזותי במוח האמצע הגבי. מבנה זה נקרא טקטום אופטי בדגים ובבעלי חוליות אחרים שאינם יונקים והוא הומולוגי לקוליקולוס העליון ביונקים3.
טקטום אופטי הוא מבנה רב שכבתי סימטרי דו-צדדי במוח האמצע הגבי. בדגי זברה וברוב הדגים האחרים, כל אונה של טקטום אופטי מקבלת קלט חזותי אך ורק מהעין הקונטרולטרלית, כך שעצב הראייה השמאלי מסתיים באונה הטקטלית הימנית ועצב הראייה הימני מסתיים באונה הטקטלית השמאלית4 (איור 1). בדומה למקבילו היונקים, הקוליקולוס המעולה, טקטום אופטי משלב מידע חזותי עם קלטים חושיים אחרים, כולל אודישן וסומטוסנסציה, שליטה בשינויים בקשב החזותי ובתנועות עיניים כגון סקאדות 1,5,6. עם זאת, שלא כמו הקוליקולוס העליון של היונקים, הטקטום האופטי מייצר ברציפות נוירונים וגליה חדשים מגומחת תאי גזע מיוחדת ליד הקצוות המדיאליים והקאודלים של האונות הטקטליות הנקראות אזור ההתרבותהטקטלית 7. תחזוקה של אבות מתרבים בטקטום האופטי ובאזורים אחרים במערכת העצבים המרכזית תורמת, בין השאר, ליכולת ההתחדשות המדהימה המתועדת בדגי זברה8.
עבודות קודמות שבחנו את מוחותיהם של דגים עיוורים או חדי עין גילו כי גודל טקטום אופטי עומד ביחס ישר לכמות העצבנות ברשתית שהוא מקבל 9,10,11. בדגי מערות בוגרים, שעיניהם מתנוונות באמבריוגנזה מוקדמת, טקטום הראייה קטן באופן ניכר מזה של דגי פני השטח הקרובים, בעלי הראייה הקרובה9. ניתן לחסום את ניוון העיניים של דגי המערות על ידי החלפת העדשה האנדוגנית בעדשה של דג פני השטח במהלך העובר. כאשר דגי מערות חדי עין אלה גדלים לבגרות, האונה הטקטלית העצבנית מכילה כ-10% יותר תאים מאשר האונה הטקטלית שאינה מוחננת9. באופן דומה, בדגי קטל זחלים שעברו טיפולים כימיים כדי ליצור עיניים בגדלים שונים בתוך אותו אדם, הצד של הטקטום עם יותר עצבנות היה גדול יותר והכיל יותר נוירונים10. עדויות מניסויים בריסוק עצבי ראייה בדגי זהב בוגרים מצביעות על כך שהתרבות העצבנות מקדמת התפשטות, כאשר התפשטות תאי הכרק פוחתת כאשר העצבנות שובשה11.
כדי לאשר ולהרחיב את המחקרים הקלאסיים האלה, כמה דיווחים עדכניים מספקים נתונים המצביעים על כך שהתפשטות בתגובה להפנמה מווסתת, לפחות בחלקה, על ידי מסלול BDNF-TrkB12,13. שאלות פתוחות רבות לגבי גדילה והתפתחות של טקטום אופטי נותרו בעינן, כולל כיצד מערכת חושית מתפתחת מתמודדת עם פציעה וניוון אקסון, אילו אותות תאיים ומולקולריים מאפשרים לקלט רשתית לווסת את צמיחת הטקטום האופטי, מתי מנגנונים אלה הופכים לפעילים, והאם התפשטות והתמיינות הקשורות לעירוי העצבנות מאפשרות לרשתית ולרקמת היעד שלה לתאם את קצבי הגדילה ולהבטיח מיפוי רטינוטופי מדויק. בנוסף, ישנן שאלות הרבה יותר גדולות על התפתחות תלוית פעילות שניתן לטפל בהן על ידי חקירת מערכת הראייה של דגי הזברה עם גישות כירורגיות כמו זו המתוארת להלן.
כדי לחקור את המנגנונים התאיים והמולקולריים שבאמצעותם פעילות עצבית, במיוחד מתוך קלט חזותי, משנה את הישרדות התאים ואת התפשטותם, הגישה המתוארת משווה באופן ישיר בין אונות טקטליות עצבוניות ומנוכרות (איור 1) בתוך זחלי דגי זברה בודדים. שיטה זו מאפשרת תיעוד של התנוונות אקסון RGC בטקטום האופטי ואישור לכך שמספר התאים המיטוטיים מתואם עם העצבנות.
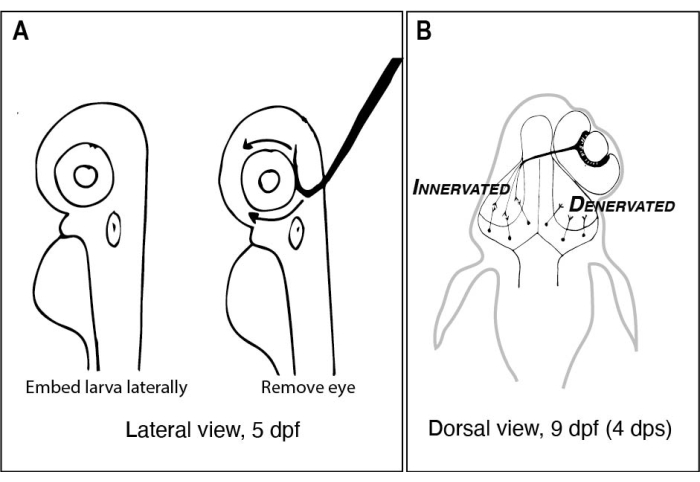
איור 1: רישומים של זחלי דגי זברה לפני ואחרי הסרת עיניים חד-צדדית. (A) ציור של 5 זחלי dpf כפי שנצפו תחת מיקרוסקופ מנתח. כל זחל משובץ באגרוז בעל נקודת התכה נמוכה ומכוון לרוחב לפני שמחט טונגסטן עם קצה חד ומחובר משמשת כדי להוציא את העין הפונה כלפי מעלה (עין שמאל בדוגמה זו). (B) ציור הנוף הגבי של זחל 9 dpf כתוצאה מהניתוח המתואר ב-A. רק שלושה אקסוני RGC שעברו מיפוי גבוה מעין ימין נראים מתייבשים ומתחברים לנוירונים באונה הטקטלית השמאלית. קיצורים: dpf = ימים לאחר ההפריה; dps = ימים לאחר הניתוח; RGC = תאי גנגליון רשתית. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
