Proteinfosforylering styr de flesta cellulära processer, inklusive men inte begränsat till svaret på DNA-skador, tillväxtfaktorsignalering och passagen genom mitos 1,2,3. I däggdjursceller fosforyleras majoriteten av proteinerna vid en eller flera serin-, treonin- eller tyrosinrester vid någon tidpunkt, med fosfoseriner och fosfotreoniner som utgör cirka 98% av alla fosforyleringsställen 2,3. Medan kinaser har studerats omfattande inom cellulär signalering, är PPP: s roll i regleringen av dynamiska cellulära processer fortfarande framväxande.
Fosforyleringsdynamiken styrs av det dynamiska samspelet mellan kinaser och fosfataser. I däggdjursceller finns det mer än 400 proteinkinaser som katalyserar serin / treoninfosforylering. Över 90% av dessa platser defosforyleras av fosfoproteinfosfataser (PPP), en liten familj av enzymer som består av PP1, PP2A, PP2B, PP4-7, PPT och PPZ 2,3. PP1 och PP2A är ansvariga för majoriteten av fosfoserin och fosfotreonin defosforylering inom en cell 2,3,4. Den anmärkningsvärda skillnaden i antal mellan kinaser och fosfataser och bristen på specificitet hos PPP-katalytiska subenheter in vitro ledde till tron att kinaser är den viktigaste determinanten för fosforylering 2,3. Emellertid, flera studier har visat fosfataser för att fastställa substratspecificitet genom bildandet av multimera holoenzymer 5,6,7,8,9. Till exempel är PP1 en heterodimer som består av en katalytisk underenhet och vid en given tidpunkt en av de mer än 150 reglerande underenheterna 6,7,8. Omvänt är PP2A en heterotrimer som bildas av en byggnadsställning (A), en reglerande (B) och en katalytisk (C) underenhet 2,3,9. Det finns fyra distinkta familjer av PP2A-regulatoriska underenheter (B55, B56, PR72 och striatin), var och en med flera gener, skarvvarianter och lokaliseringsmönster 2,3,9. Offentlig-privata partnerskapens multimeriska karaktär fyller gapet i antalet kinaser och PPP-katalytiska underenheter. Det skapar dock analytiska utmaningar för att studera PPP-signalering. För att omfattande analysera PPP-signalering är det viktigt att undersöka de olika holoenzymerna i en cell eller vävnad. Stora framsteg har gjorts för att studera det mänskliga släktomet genom användning av kinashämmare pärlor, kallade multiplexhämmare pärlor eller kinobeads, en kemisk proteomisk strategi där kinashämmare immobiliseras på pärlor och masspektrometri används för att identifiera berikade kinaser och deras interagerare 10,11,12,13.
Vi har etablerat ett liknande tillvägagångssätt för att studera PPP-biologi. Denna teknik involverar affinitetsfångst av PPP-katalytiska underenheter med användning av pärlor med en immobiliserad, icke-selektiv PPP-hämmare som kallas mikrocystin-LR (MCLR) som kallas fosfatashämmarepärlor (PIBs)14,15. Till skillnad från andra metoder som kräver endogen märkning eller uttryck av exogena PPP-underenheter som kan förändra proteinaktivitet eller lokalisering, möjliggör PIB-MS anrikning av endogena PPP-katalytiska underenheter, deras associerade reglerande och byggnadsställningar och interagerande proteiner (kallat PPPome) från celler och vävnader vid en given tidpunkt eller under specifika behandlingsförhållanden. MCLR hämmar PP1, PP2A, PP4-6, PPT och PPZ vid nanomolära koncentrationer, vilket gör PIBs mycket effektiva vid anrikning för PPPome16. Denna metod kan skalas för användning på vilket utgångsmaterial som helst från celler till kliniska prover. Här beskriver vi i detalj användningen av PIB och masspektrometri (PIB-MS) för att effektivt fånga, identifiera och kvantifiera det endogena PPPome och dess modifieringstillstånd.
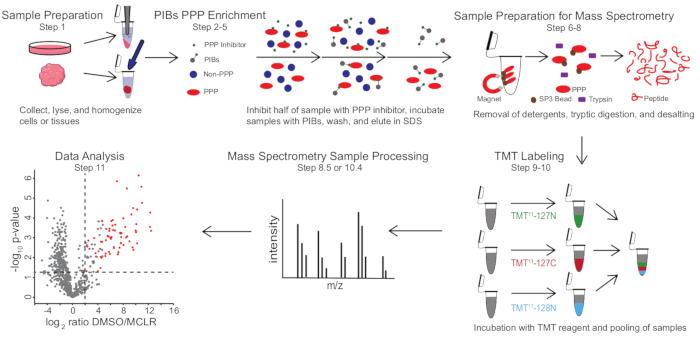
Bild 1: Visuell sammanfattning av PIB-MS-protokollet. I ett PIB-MS-experiment kan prover erhållas i olika former, från celler till tumörer. Provet samlas in, lyseras och homogeniseras före PPP-anrikning. För att berika för PPP inkuberas lysaten med PIBs med eller utan en PPP-hämmare, såsom MCLR. PDB tvättas sedan och växtskyddsmedel elueras under denatureringsförhållanden. Proverna bereds för masspektrometrianalys genom avlägsnande av tvättmedel genom SP3-proteinanrikning, tryptisk matsmältning och avsaltning. Prover kan sedan valfritt TMT-märkas före masspektrometrianalys. Klicka här för att se en större version av denna siffra.
PIB-MS involverar lys och förtydligande av celler eller vävnader, inkubation av lysaten med PIBs, eluering och analys av eluatet via western blotting eller masspektrometribaserade metoder (Figur 1). Tillägget av fri MCLR kan användas som en kontroll för att skilja specifika PIB-bindemedel från icke-specifika interagerare. För de flesta applikationer kan ett etikettfritt tillvägagångssätt användas för att direkt identifiera proteiner i eluater. I de fall där större precision i kvantifieringen eller identifieringen av arter med låg förekomst behövs, kan ytterligare bearbetning med tandem-masstagg (TMT) -märkning användas för att öka täckningen och minska inmatningen.
