Den menneskelige krop har anslået 10-100 billioner levende mikrobielle celler (bakterier, arkæa svampe), som primært findes i tarm-, hud- og slimhindemiljøerne1. I en sund tilstand giver disse fordele for deres vært, herunder vitaminproduktion, modning af immunsystemet, stimulering af medfødte og adaptive immunresponser på patogener, regulering af fedtstofskifte, modulering af stressresponser og mere med indvirkning på vækst og udvikling, sygdomsdebut og aldring 2,3,4,5 . Tarmens mikrobiota udvikler sig også betydeligt gennem hele livet. Den mest drastiske udvikling forekommer i barndommen og den tidlige barndom6, men betydelige ændringer forekommer også med alderen, herunder et fald i overflod af Bifidobacterium og en stigning i Clostridium, Lactobacillus, Enterobacteriaceae og Enterococcus arter7. Livsstil kan yderligere ændre tarmens mikrobielle sammensætning, hvilket fører til dysbiose (tab af gavnlige bakterier, overvækst af opportunistiske bakterier), hvilket resulterer i forskellige patologier såsom inflammatorisk tarmsygdom, diabetes og fedme5, men også bidrager til Alzheimers og Parkinsons sygdomme 8,9,10,11.
Denne erkendelse har kritisk bidraget til at forfine begrebet tarm-hjerneakse (GBA), hvor interaktioner mellem tarmfysiologi (nu inklusive mikroberne i den) og nervesystemet betragtes som den vigtigste regulator for dyremetabolisme og fysiologiske funktioner12. Imidlertid er mikrobiotaens præcise rolle i tarm-hjerne-signalering og de tilhørende virkningsmekanismer langt fra fuldt ud forstået13. Da tarmmikrobiota er en vigtig determinant for sund aldring, er hvordan bakterier modulerer aldringsprocessen blevet genstand for intens forskning og kontrovers 6,14,15.
Med demonstrationen af, at rundorm Caenorhabditis elegans er vært for en bonafide tarmmikrobiota domineret – som i andre arter – af Bacteroidetes, Firmicutes og Actinobacteria 16,17,18,19,20, dens hurtige stigning som en eksperimentel platform til at studere vært-tarm kommensale interaktioner 21,22,23,24 ,25,26 har udvidet vores efterforskningsarsenal betydeligt26,27,28,29. Især kan eksperimentelle tilgange med høj kapacitet, der er tilgængelige for C. elegans til at studere gen-diæt, gen-lægemiddel, genpatogen osv. interaktioner, tilpasses til hurtigt at undersøge, hvordan bakterieisolater og cocktails påvirker C. elegans sundhed og aldring.
Den nuværende protokol beskriver en eksperimentel pipeline til straks at screene arrays af bakterielle isolater eller blandinger sat i multiwell plader for virkninger på C. elegans stressresistens som en proxy for sundhed, som kan bruges til at identificere probiotika. Den beskriver, hvordan man dyrker store ormepopulationer og håndterer bakterielle arrays i 96- og 384-brønds pladeformater, før orme behandles til automatiseret stressresistensanalyse ved hjælp af en fluorescenspladelæser (figur 1). Tilgangen er baseret på etiketfri automatiserede overlevelsesanalyser (LFASS)30 , der udnytter fænomenet dødsfluorescens31, hvorved døende orme producerer et udbrud af blå fluorescens, der kan bruges til at lokalisere dødstidspunktet. Blå fluorescens udsendes af glucosylestere af anthranilsyre opbevaret i C. elegans tarmgranulat (en type lysosomrelateret organel), som brister, når en nekrotisk kaskade udløses i ormens tarm ved død31.
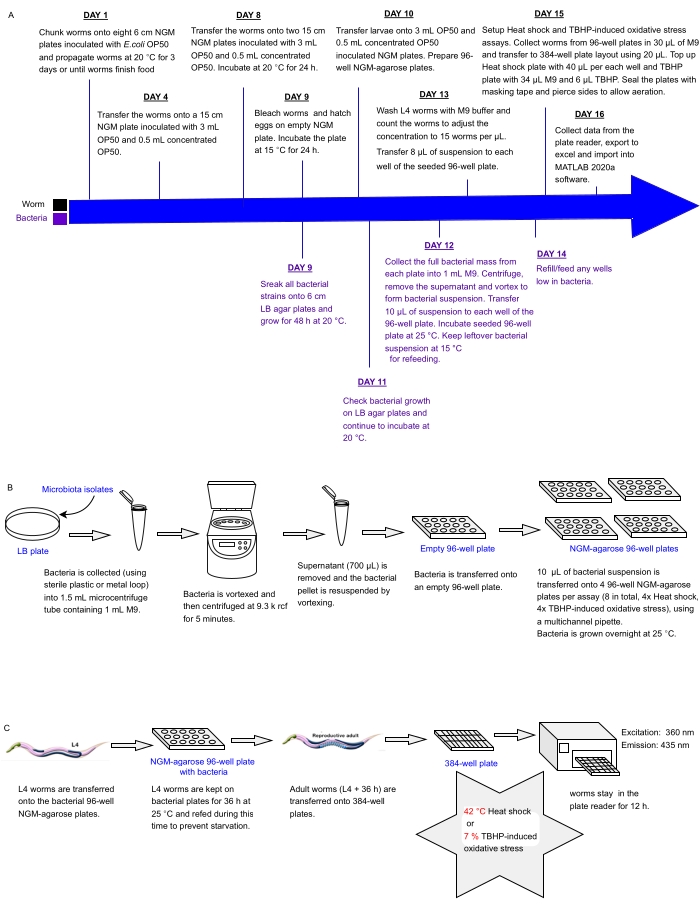
Figur 1: Eksperimentel arbejdsgang for high-throughput screening af bakterieisolater med indvirkning på C. elegans modstandsdygtighed over for stress . (A) Tidslinje for orm og bakteriel vedligeholdelse og assay opsætning. (B) 96-brønds bakteriel pladearray opsætning og håndtering. (C) opsætning af 384-brønds ormplade. Klik her for at se en større version af denne figur.
