Het menselijk lichaam herbergt naar schatting 10-100 biljoen levende microbiële cellen (bacteriën, archaea-schimmels), die voornamelijk worden aangetroffen in de darm-, huid- en mucosale omgevingen1. In een gezonde toestand bieden deze voordelen voor hun gastheer, waaronder vitamineproductie, rijping van het immuunsysteem, stimulatie van aangeboren en adaptieve immuunresponsen op pathogenen, regulatie van vetmetabolisme, modulatie van stressreacties en meer, met een impact op groei en ontwikkeling, het begin van de ziekte en veroudering 2,3,4,5 . De darmmicrobiota evolueert ook aanzienlijk gedurende het hele leven. De meest drastische evolutie vindt plaats tijdens de kindertijd en de vroege kindertijd6, maar significante veranderingen treden ook op met de leeftijd, waaronder een afname van de overvloed aan Bifidobacterium en een toename van Clostridium, Lactobacillus, Enterobacteriaceae pt Enterococcus species7. Levensstijl kan de microbiële samenstelling van de darm verder veranderen, wat leidt tot dysbiose (verlies van nuttige bacteriën, overgroei van opportunistische bacteriën), wat resulteert in verschillende pathologieën zoals inflammatoire darmaandoeningen, diabetes en obesitas5, maar ook bijdragen aan de ziekte van Alzheimer en Parkinson 8,9,10,11.
Dit besef heeft kritisch bijgedragen aan het verfijnen van het concept van de darm-hersenas (GBA), waar interacties tussen darmfysiologie (nu inclusief de microben erin) en het zenuwstelsel worden beschouwd als de belangrijkste regulator van dierlijk metabolisme en fysiologische functies12. De precieze rol van microbiota in darm-hersensignalering en de bijbehorende werkingsmechanismen zijn echter nog lang niet volledig begrepen13. Met darmmicrobiota als een belangrijke determinant van gezond ouder worden, is hoe bacteriën het verouderingsproces moduleren een onderwerp geworden van intensief onderzoek en controverse 6,14,15.
Met de demonstratie dat de rondworm Caenorhabditis elegans een bonafide darmmicrobiota herbergt die wordt gedomineerd – zoals bij andere soorten – door Bacteroidetes, Firmicutes pt Actinobacteria 16,17,18,19,20, is de snelle opkomst ervan als een experimenteel platform om gastheer-darm commensale interacties te bestuderen 21,22,23,24 ,25,26 heeft ons onderzoeksarsenaal aanzienlijk uitgebreidmet 26,27,28,29. In het bijzonder kunnen high-throughput experimentele benaderingen die beschikbaar zijn voor C. elegans om gen-dieet, gen-medicijn, gen-pathogeen, enz. interacties te bestuderen, worden aangepast om snel te onderzoeken hoe bacteriële isolaten en cocktails de gezondheid en veroudering van C. elegans beïnvloeden.
Het huidige protocol beschrijft een experimentele pijplijn om in één keer arrays van bacteriële isolaten of mengsels in multiwell-platen te screenen op effecten op C. elegans stressresistentie als een proxy voor gezondheid, die kan worden gebruikt om probiotica te identificeren. Het beschrijft hoe grote wormpopulaties kunnen groeien en bacteriële arrays in 96- en 384-well plaatformaten kunnen worden verwerkt voordat wormen worden verwerkt voor geautomatiseerde stressbestendigheidsanalyse met behulp van een fluorescentieplaatlezer (figuur 1). De aanpak is gebaseerd op labelvrije geautomatiseerde overlevingstests (LFASS)30 die gebruik maken van het fenomeen van doodsfluorescentie31, waarbij stervende wormen een uitbarsting van blauwe fluorescentie produceren die kan worden gebruikt om het tijdstip van overlijden te bepalen. Blauwe fluorescentie wordt uitgestoten door glucosylesters van anthranilic acid opgeslagen in C. elegans darmkorrels (een soort lysosoom-gerelateerd organel), die barsten wanneer een necrotische cascade wordt geactiveerd in de worm darm bij overlijden31.
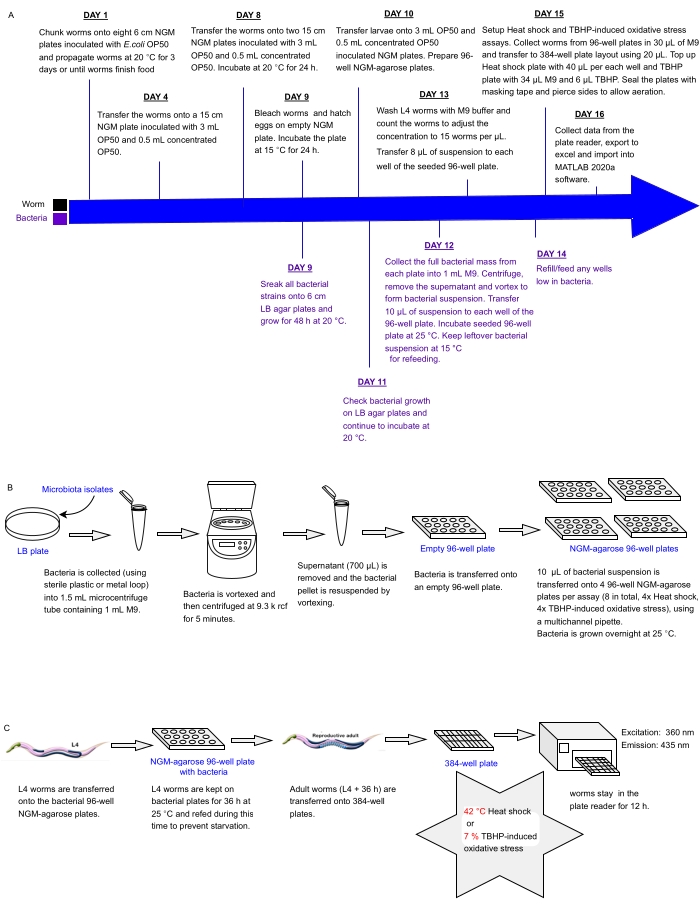
Figuur 1: Experimentele workflow voor high-throughput screening van bacteriële isolaten met impact op C. elegans resistentie tegen stress. (A) Tijdlijn voor worm- en bacterieonderhoud en testopstelling. (B) 96-well bacteriële plaat array setup en handling. (C) 384-well wormplaat opstelling. Klik hier om een grotere versie van deze figuur te bekijken.
