Menneskekroppen har anslagsvis 10-100 billioner levende mikrobielle celler (bakterier, archaea sopp), som primært finnes i tarm-, hud- og slimhinnemiljøer1. I en sunn tilstand gir disse fordeler for verten, inkludert vitaminproduksjon, modning av immunsystemet, stimulering av medfødte og adaptive immunresponser mot patogener, regulering av fettmetabolismen, modulering av stressresponser og mer, med innvirkning på vekst og utvikling, sykdomsutbrudd og aldring 2,3,4,5 . Tarmmikrobiotaen utvikler seg også betydelig gjennom livet. Den mest drastiske utviklingen skjer i barndom og tidlig barndom6, men signifikante endringer forekommer også med alderen, inkludert en reduksjon i Bifidobacterium overflod og en økning i Clostridium, Lactobacillus, Enterobacteriaceae og Enterococcus arter7. Livsstil kan ytterligere endre tarmens mikrobielle sammensetning som fører til dysbiose (tap av gunstige bakterier, overvekst av opportunistiske bakterier), noe som resulterer i ulike patologier som inflammatorisk tarmsykdom, diabetes og fedme5, men bidrar også til Alzheimers og Parkinsons sykdommer 8,9,10,11.
Denne erkjennelsen har kritisk bidratt til å raffinere begrepet tarm-hjerneaksen (GBA), hvor interaksjoner mellom tarmfysiologi (nå inkludert mikrober i den) og nervesystemet regnes som hovedregulator for dyremetabolisme og fysiologiske funksjoner12. Imidlertid er den nøyaktige rollen som mikrobiota i tarm-hjernesignalering og tilhørende virkningsmekanismer langt fra å bli fullt ut forstått13. Med tarmmikrobiota som en viktig determinant for sunn aldring, har hvordan bakterier modulerer aldringsprosessen blitt gjenstand for intens forskning og kontrovers 6,14,15.
Med demonstrasjonen at rundorm Caenorhabditis elegans er vert for en bonafide tarmmikrobiota dominert – som i andre arter – av Bacteroidetes, Firmicutes og Actinobacteria 16,17,18,19,20, den raske økningen som en eksperimentell plattform for å studere host-gut commensal interaksjoner 21,22,23,24 ,25,26 har betydelig utvidet vårt etterforskningsarsenal26,27,28,29. Spesielt kan eksperimentelle tilnærminger med høy gjennomstrømning som er tilgjengelige for C. elegans for å studere gen-diett, gen-stoff, genpatogen, etc. interaksjoner, tilpasses for raskt å undersøke hvordan bakterielle isolater og cocktailer påvirker C. elegans helse og aldring.
Den nåværende protokollen beskriver en eksperimentell rørledning for å skjerme samtidig arrays av bakterielle isolater eller blandinger satt i multiwell plater for effekter på C. elegans stressmotstand som en proxy for helse, som kan brukes til å identifisere probiotika. Den beskriver hvordan man kan vokse store ormpopulasjoner og håndtere bakterielle arrays i 96- og 384-brønns plateformater før behandling av ormer for automatisert stressmotstandsanalyse ved hjelp av en fluorescensplateleser (figur 1). Tilnærmingen er basert på etikettfrie automatiserte overlevelsesanalyser (LFASS)30 som utnytter fenomenet dødsfluorescens31, der døende ormer produserer et utbrudd av blå fluorescens som kan brukes til å fastslå dødstidspunktet. Blå fluorescens sendes ut av glukosylestere av anthranilinsyre lagret i C. elegans tarmgranulat (en type lysosomrelatert organell), som brister når en nekrotisk kaskade utløses i ormtarmen ved døden31.
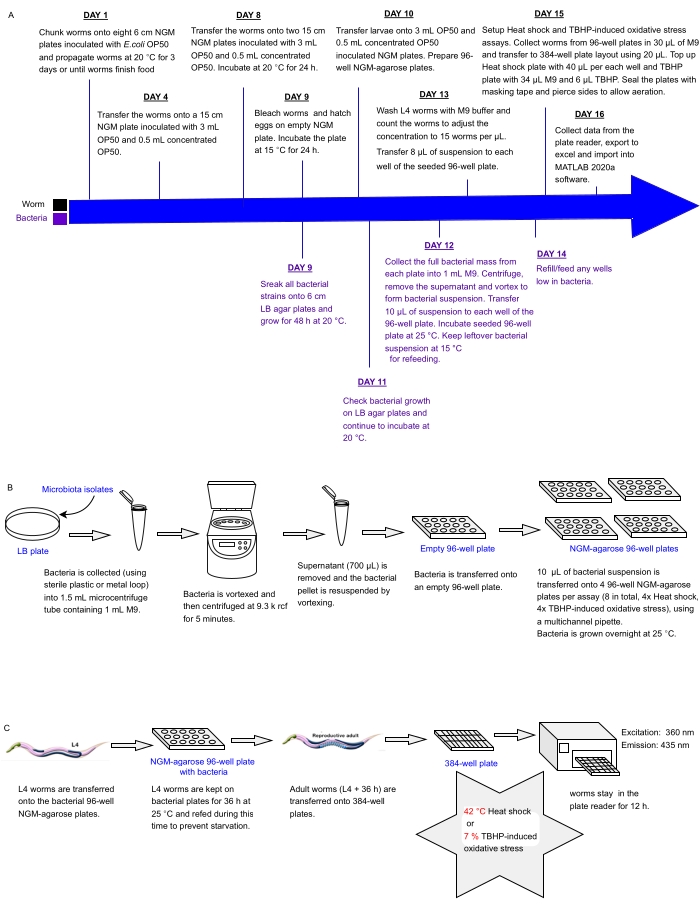
Figur 1: Eksperimentell arbeidsflyt for høy gjennomstrømningsscreening av bakterieisolater med innvirkning på C. elegans motstand mot stress . (A) Tidslinje for vedlikehold og analyse av ormer og bakterier. (B) 96-brønn bakteriell plate array oppsett og håndtering. (C) 384-brønns ormplateoppsett. Klikk her for å se en større versjon av dette tallet.
