Человеческое тело содержит около 10-100 триллионов живых микробных клеток (бактерий, грибов архей), которые в основном находятся в кишечнике, коже и слизистой оболочке1. В здоровом состоянии они обеспечивают преимущества для своего хозяина, включая выработку витаминов, созревание иммунной системы, стимуляцию врожденных и адаптивных иммунных реакций на патогены, регуляцию жирового обмена, модуляцию стрессовых реакций и многое другое, с влиянием на рост и развитие, начало заболевания и старение 2,3,4,5 . Микробиота кишечника также значительно развивается на протяжении всей жизни. Наиболее радикальная эволюция происходит в младенчестве и раннем детстве6, но значительные изменения также происходят с возрастом, включая снижение численности бифидобактерий и увеличение Clostridium, Lactobacillus, Enterobacteriaceae и Enterococcus видов 7. Образ жизни может еще больше изменить микробный состав кишечника, что приводит к дисбактериозу (потеря полезных бактерий, чрезмерный рост условно-патогенных бактерий), что приводит к различным патологиям, таким как воспалительные заболевания кишечника, диабет и ожирение5, но также способствует развитию болезней Альцгеймера и Паркинсона 8,9,10,11.
Это осознание критически способствовало уточнению концепции оси кишечник-мозг (GBA), где взаимодействия между физиологией кишечника (теперь включая микробы внутри него) и нервной системой считаются основным регулятором метаболизма животных и физиологических функций12. Однако точная роль микробиоты в передаче сигналов кишечником и мозгом и связанные с ней механизмы действия далеко не полностью поняты13. Поскольку микробиота кишечника является ключевым фактором, определяющим здоровое старение, то, как бактерии модулируют процесс старения, стало предметом интенсивных исследований и споров 6,14,15.
С демонстрацией того, что круглый червь Caenorhabditis elegans содержит добросовестную микробиоту кишечника, в которой преобладают, как и у других видов, Bacteroidetes, Firmicutes и Actinobacteria 16,17,18,19,20, его быстрый рост в качестве экспериментальной платформы для изучения комменсальных взаимодействий между кишечником хозяина и кишечника 21,22,23,24 ,25,26 значительно расширил наш следственный арсенал 26,27,28,29. В частности, высокопроизводительные экспериментальные подходы, доступные для C. elegans для изучения взаимодействий ген-диета, ген-лекарство, ген-патоген и т. Д., Могут быть адаптированы для быстрого изучения того, как бактериальные изоляты и коктейли влияют на здоровье и старение C. elegans.
Настоящий протокол описывает экспериментальный конвейер для одновременного скрининга массивов бактериальных изолятов или смесей, установленных в многолуночных пластинах, на предмет воздействия на стрессоустойчивость C. elegans в качестве прокси для здоровья, который может быть использован для идентификации пробиотиков. В нем подробно описывается, как выращивать большие популяции червей и обрабатывать бактериальные массивы в форматах пластин с 96 и 384 лунками перед обработкой червей для автоматизированного анализа стрессоустойчивости с использованием считывателя флуоресцентных пластин (рисунок 1). Этот подход основан на автоматизированных анализах выживания без маркировки (LFASS)30 , которые используют явление флуоресценциисмерти 31, в результате чего умирающие черви производят всплеск синей флуоресценции, который может быть использован для точного определения времени смерти. Синяя флуоресценция испускается глюкозильными эфирами антраниловой кислоты, хранящимися в кишечных гранулах C. elegans (тип органелл, связанных с лизосомами), которые лопаются, когда некротический каскад запускается в кишечнике червя после смерти31.
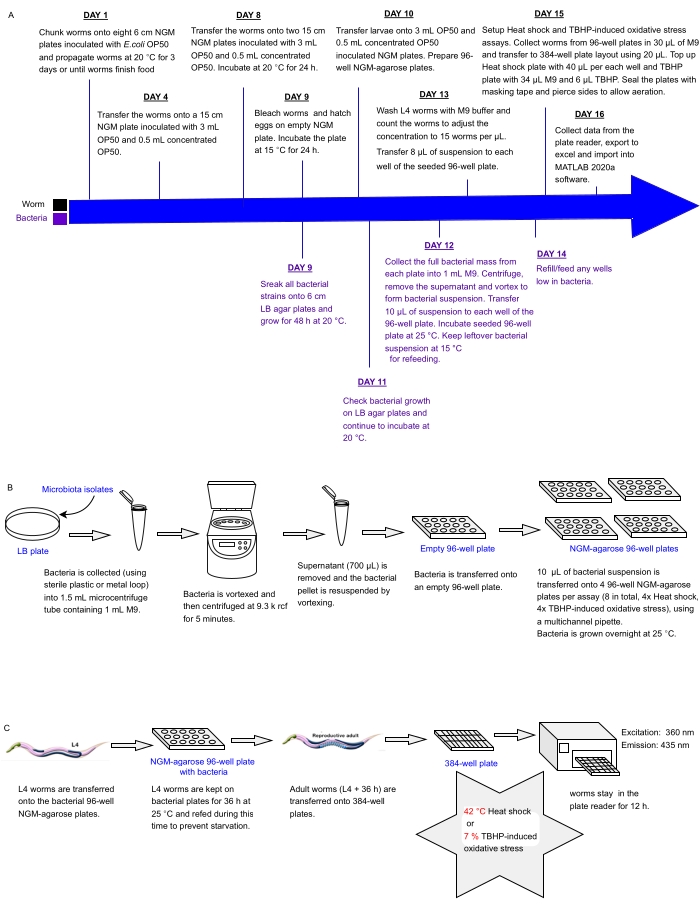
Рисунок 1: Экспериментальный рабочий процесс для высокопроизводительного скрининга бактериальных изолятов с воздействием на устойчивость C. elegans к стрессу. (A) Временная шкала для поддержания червей и бактерий и настройки анализа. (B) Установка и обработка 96-луночной бактериальной пластинчатой решетки. (C) Установка червячной пластины на 384 скважины. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
