D. melanogaster animals expressing red light-sensitive opsins eNpHR2.0 or ReaChR in the heart tube were generated by obtaining progeny from the cross between each UAS-opsin transgenic line and Hand-GAL4 driver. The tissue specificity of the GAL4 driver was verified by imaging GFP expression (Figure 4). Drosophila 3rd instar larva and early pupa developmental stages were used to demonstrate the effects of eNpHR2.0 and ReaChR activation by red light. Designed ~617 nm red-light pulses, delivered by LED, illuminated the larva/pupa and activated the eNpHR2.0 and ReaChR in the heart. Although the reported maximum response wavelength of NpHR is ~580 nm and of ReaChR is ~600 nm, 617 nm light illumination can penetrate deeper with enhanced light energy delivery toward the opsin-expressing heart tissue22.
Mounted on the microscope slide with the dorsal side down in the inverted microscope setup, the larva/pupa was illuminated by an LED light beam directed to the A7 body segment. Examples of the body cross-section images are shown in Figure 5A and Figure 6A. The heart appears as a contracting and dilating circular shape in the video recordings consisting of 4,000 frames (Supplementary videos 1–6). To mimic different heart conditions, four types of light pulses were designed. A single pulse lasting 10 s after 5 s waiting time generated restorable heart arrest induced by eNpHR2.0, as shown in Figure 5B. For the heart pacing at frequencies slower than the resting heart rate (RHR), mediated by eNpHR2.0, two light-pulse sequences with pacing frequencies equal to RHR/2 and RHR/4 lasting 8 s with a waiting time of 6 s in between were used (Figure 5C). The duty cycle of each light pulse sequence was 90%. This light stimulation regimen caused a heart condition reminiscent of bradycardia. The stimulation pattern to increase the heart rate due to ReaChR activation consisted of three sequences of light pulses at frequencies of RHR + 0.5 Hz, RHR + 1 Hz, and RHR + 1.5 Hz, respectively, with a pulse width of 20 ms (Figure 5D). This pulse regimen was aimed at causing a tachycardic heart condition. The light power density was 7.49 mW/mm2 during all the experiments. For control experiments, no light illumination was set.
Each experimental variant was recorded five times. M-mode videos of the fly heart were processed into 2D masks using FlyNet 2.027. This software automatically segments the heart region to produce the cardiac function data sets. The program provides a mask of the heart in each frame, which can be further corrected manually, if needed, to generate accurate quantification of the functional parameters of the beating heart, such as heart rate (HR), end-diastolic dimension (EDD), and end-systolic dimension (ESD), fractional shortening (FR), end-diastolic area (EDA), end-systolic area (ESA), etc. The heart rate is measured by analyzing the heart area over time. The control video with no light pulses is used to establish a baseline heart rate (e.g., RHR) for each animal.
Figure 5B and Figure 6B show 10 s long heart arrest caused by Hand>eNpHR2.0 activation using red light (617 nm) in larva and pupa, respectively. When the red light was turned on, the Drosophila's heart stopped beating and remained in this state until the end of the light illumination. The heart function was restored after the red light was turned off. Animals that did not have opsin expressed ("no opsin" control) did not respond to the red-light illumination (Supplementary Figure 2A and Supplementary Figure 3A). The control experiments with Hand>eNpHR2.0 animals where the 10 s red light illumination was not turned on ("no light" control) showed the heart beating normally (Supplementary Figure 4A and Supplementary Figure 4C).
Using Hand>eNpHR2.0 animals, red-light pulses at frequencies lower than the RHR were applied. The heart contraction frequency was reduced following the light signals; this slower heart rate mimics one type of heart arrhythmia, bradycardia (Figure 5C and Figure 6C for larva and pupa, respectively). The slower heart pacing was not observed in "no opsin" (Supplementary Figure 2B and Supplementary Figure 3B) and in "no light" (Supplementary Figure 4A and Supplementary Figure 4C) control experiments.
Increasing the heart rates can be achieved by activating Hand>ReaChR opsin with red-light pulse trains at a frequency higher than the RHR of the given animal. A series of three light pulse trains at different stimulation frequencies (e.g., RHR + 0.5 Hz, RHR + 1 Hz, RHR + 1.5 Hz) were applied on Hand>ReaChR larvae and pupae hearts. The obtained data clearly shows increased heart rate following the light pulses (Figure 5D and Figure 6D for larva and pupa, respectively). The heart condition demonstrated in these experiments mimics tachycardia. Negative control experiments are shown in Supplementary Figure 2C, Supplementary Figure 3C, and Supplementary Figure 4B,D.
Overall, the results demonstrate the feasibility of non-invasive and specific optogenetic control of the heart rhythm at various developmental stages in transgenic animal models of D. melanogaster.
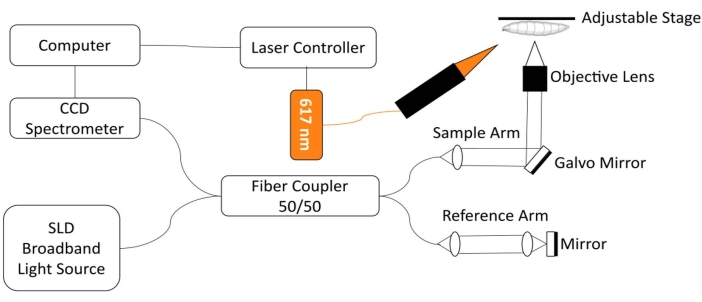
Figure 1: OCT imaging system integrated with 617 nm LED module for optogenetic control of Drosophila heart function. Please click here to view a larger version of this figure.
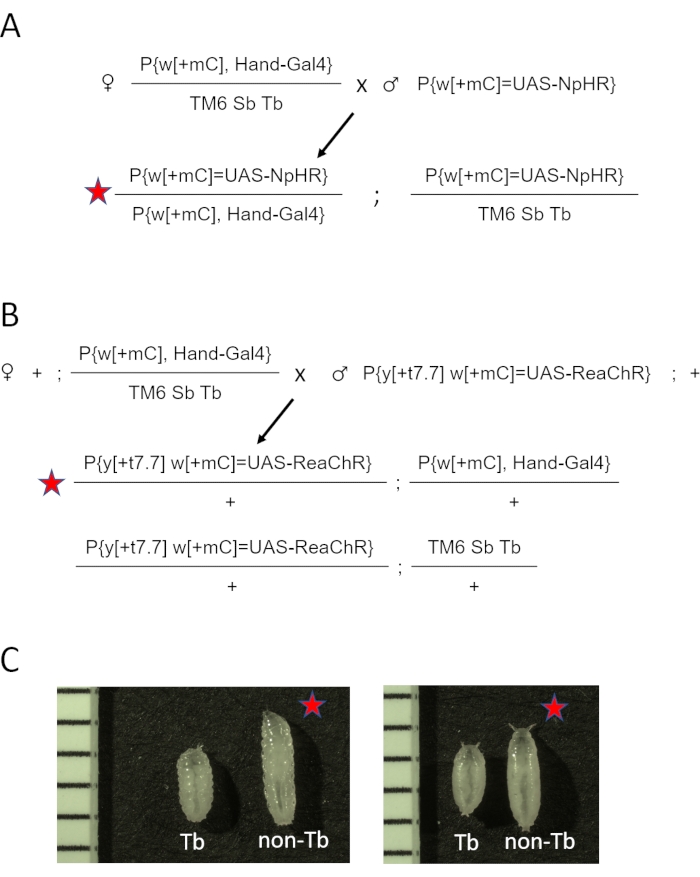
Figure 2: Generating D. melanogaster animals expressing opsin in the heart. (A) Genetic cross diagram. Females Hand-GAL4/TM6 SbTb were crossed to males carrying eNpHR2.0. The resulting Hand-GAL4/eNpHR2.0 progeny (marked by the red star) were collected for OCT imaging, and Hand-GAL4/TM6 Sb Tb were discarded based on their phenotypic appearance. (B) Genetic cross diagram. Females Hand-GAL4/TM6 SbTb were crossed to males carrying ReaChR. The resulting Hand-GAL4/ReaChR progeny (marked by the red star) were collected for OCT imaging, and Hand-GAL4/TM6 Sb Tb were discarded based on their phenotypic appearance. (C) Phenotypic differences between Hand-GAL4/opsin (red star) and Hand-GAL4/TM6 Tb progeny. Animals carrying the Tb gene mutation on the TM6 chromosome have a "tubby" body shape compared to normal, non-Tb larva or pupa. The left panel shows larvae; the right panel shows early pupae. Images also include a ruler with 1 mm marks. Please click here to view a larger version of this figure.
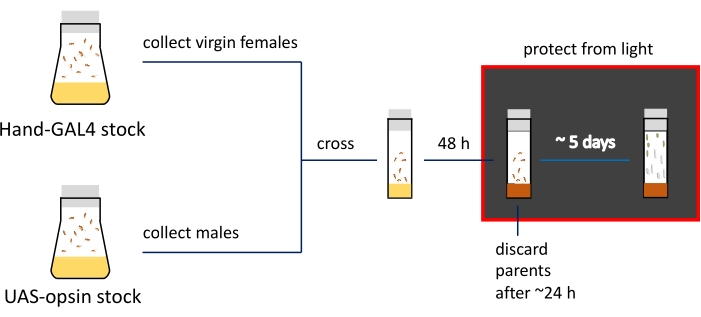
Figure 3: Schematic presentation and timeline of the imaging preparation procedures. Parental stocks are kept in fly bottles; virgin females and males are crossed in narrow vials filled with regular food (indicated by yellow color). Actively egg-laying flies are transferred to ATR-containing media (shown in brown) vials. Vials with developing progeny need to be kept in the dark from this step. 3rd instar larvae and early pupa are collected from the vial walls for imaging. Please click here to view a larger version of this figure.
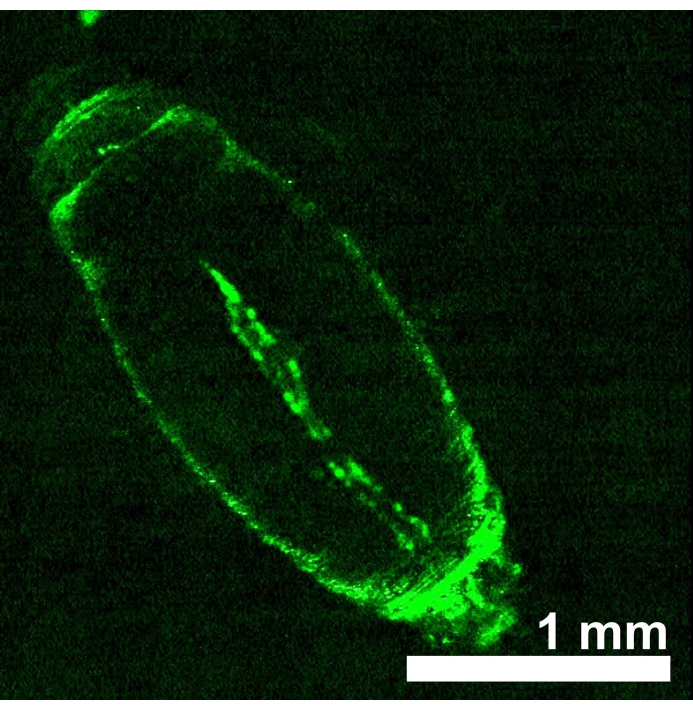
Figure 4: D. melanogaster early pupa expressing UAS-GFP (BDSC 6658) driven by Hand-GAL4 (BDSC 48396). The fluorescence pattern confirms the heart specificity of the Hand-GAL4 driver. Please click here to view a larger version of this figure.
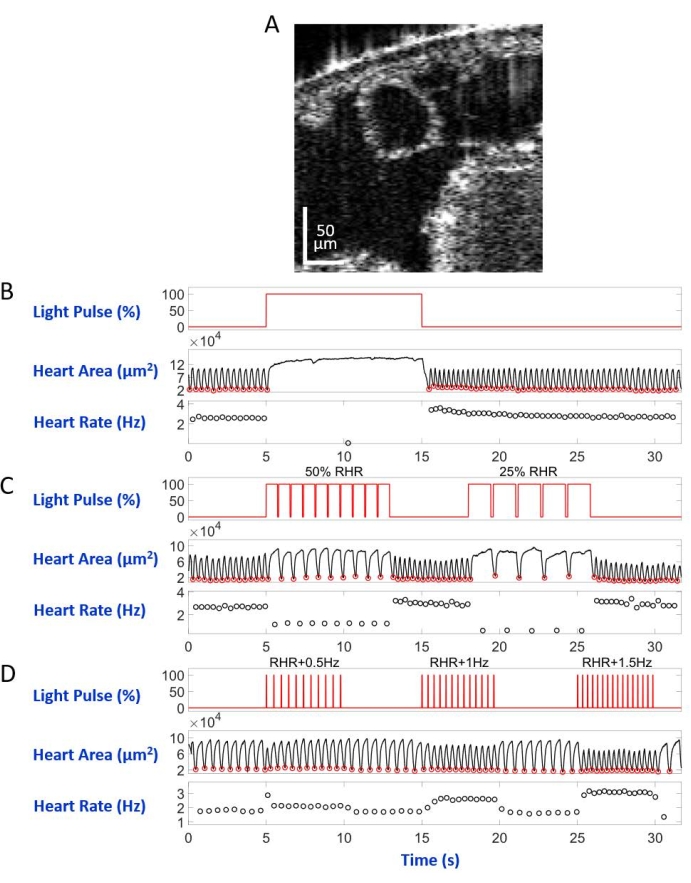
Figure 5: Simulation of heart arrest, bradycardia, and tachycardia in D. melanogaster larva. (A) OCT image of a larval body cross-section. The heart appears as a circle below the body's surface. (B) Graphic presentation of the restorable heart arrest. The upper panel shows the timing (X-axis) of the red-light illumination (Y-axis, light source power level percentage). The middle panel indicates the change in heart area (Y-axis, square micrometers) over time (X-axis). The lower panel shows the heart rate change (Y-axis, hertz) over time (X-axis). (C) Graphic presentation of eNpHR2.0-mediated restorable bradycardia. The upper panel shows pulses of the red-light illumination, inducing two periods of bradycardia: 50% of the RHR and 25% of the RHR. Heart area and heart rate changes are shown on the middle and lower panels, respectively. (D) Graphic presentation of the heart pacing by activated ReaChR. The upper panel shows a series of 20 ms red light pulses occurring at RHR + 0.5 Hz, RHR + 1 Hz, and RHR + 1.5 Hz frequencies. The heart contractions follow the light pulse frequencies, as shown on the middle and lower panels. Please click here to view a larger version of this figure.
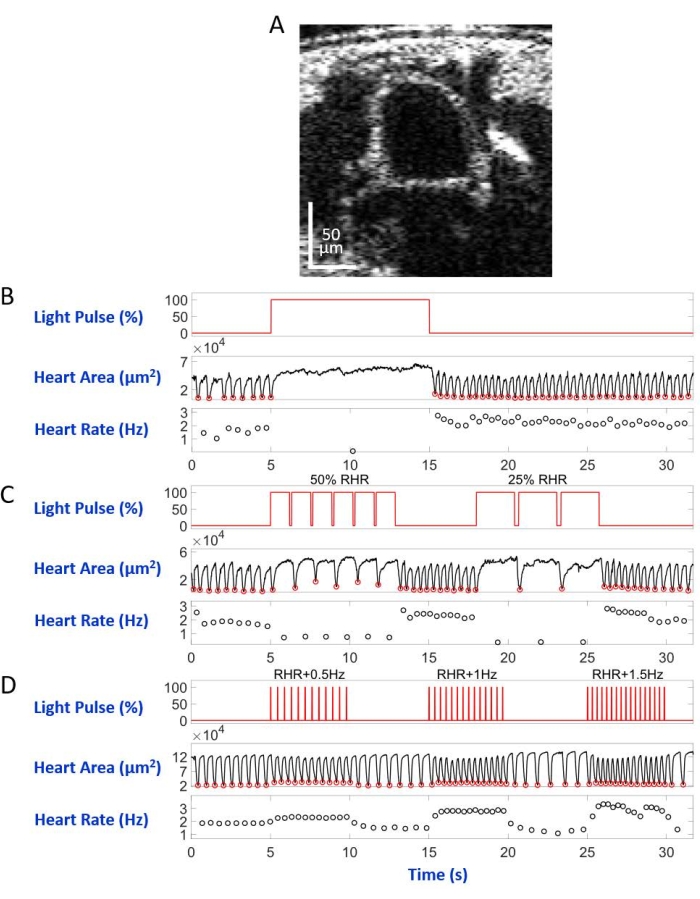
Figure 6: Simulation of heart arrest, bradycardia, and tachycardia in D. melanogaster pupa. (A) OCT image of pupal body cross-section. The heart appears as a circle below the body's surface. (B) Graphic presentation of the restorable heart arrest. The upper panel shows the timing (X-axis) of the red-light illumination (Y-axis, light source power level percentage). The middle panel indicates the change in heart area (Y-axis, square micrometers) over time (X-axis). The lower panel shows the heart rate change (Y-axis, hertz) over time (X-axis). (C) Graphic presentation of eNpHR2.0-mediated restorable bradycardia. The upper panel shows pulses of the red-light illumination, inducing two periods of bradycardia: 50% of the RHR and 25% of the RHR. The middle and lower panels show heart area and heart rate changes, respectively. (D) Graphic presentation of the heart pacing by activated ReaChR. The upper panel shows a series of 20 ms red light pulses at RHR + 0.5 Hz, RHR + 1 Hz, and RHR + 1.5 Hz frequencies. The heart contractions follow the frequencies of the light pulse, as shown on the middle and lower panels. Please click here to view a larger version of this figure.
Supplementary Figure 1: Genetic crosses to replace TM3 Sb balancer chromosome with TM6 Sb Tb. Virgin females Hand-GAL4 w+/ TM3 Sb were crossed with nub-GAL4NP3537 tub-GAL80ts w+/ TM6 Sb Tb males. Hand-GAL4 w+/ TM6 Sb Tb progeny, including virgin females and males, were selected (screening for pigmented eyes combined with tubby body shape). Selected flies were self-crossed to establish a stable stock. Please click here to download this File.
Supplementary Figure 2: In control experiments, the wild-type (wt) larva's heart rhythm does not change upon red light illumination. (A) No heart arrest was observed during the red-light illumination in wt larva. The upper panel shows the M-mode heart images. The red line indicates the illumination timing. The middle and lower panels show the heart area and the heart rates during the 32 s imaging time. (B,C) Red light pulses do not change the heart rates in wt larva. The upper panels show the M-mode heart images. The red line indicates the illumination timing. The middle and lower panels show the heart area and the heart rates during the 32 s imaging time. Please click here to download this File.
Supplementary Figure 3: In control experiments, the wild-type (wt) pupa's heart rhythm does not change upon red light illumination. (A) No heart arrest was observed during the red-light illumination in wt larva. The upper panel shows the M-mode heart images. The red line indicates the illumination timing. The middle and lower panels show the heart area and the heart rates during the 32 s imaging time. (B,C) Red light pulses do not change the heart rates in the wt pupa. The upper panels show the M-mode heart images. The red line indicates the illumination timing. The middle and lower panels show the heart area and heart rates during the 32 s imaging time. Please click here to download this File.
Supplementary Figure 4: D. melanogaster larvae and pupae expressing Hand>eNpHR2.0 or Hand>ReaChR do not show significant HR changes during OCT imaging without red light illumination. (A) The heart rates of Hand>eNpHR2.0 larva. (B) The heart rates of Hand>ReaChR larva. (C) The heart rates of Hand>eNpHR2.0 pupa. (D) The heart rates of Hand>ReaChR pupa. Please click here to download this File.
Supplementary Video 1: Activated eNpHR2.0 causes heart arrest in D. melanogaster larva. Please click here to download this Video.
Supplementary Video 2: Activated eNpHR2.0 causes heart arrest in D. melanogaster pupa. Please click here to download this Video.
Supplementary Video 3: eNpHR2.0-mediated restorable bradycardia in D. melanogaster larva. Please click here to download this Video.
Supplementary Video 4: eNpHR2.0-mediated restorable bradycardia in D. melanogaster pupa. Please click here to download this Video.
Supplementary Video 5: Heart pacing by activated ReaChR in D. melanogaster larva. Please click here to download this Video.
Supplementary Video 6: Heart pacing by activated ReaChR in D. melanogaster pupa. Please click here to download this Video.
