有无数的模型可用于研究癌症的进展,每个模型都是独一无二的,代表了这种复杂疾病的一种亚型。每个模型都为癌症生物学提供了独特而有价值的见解,并改进了模拟实际疾病状况的方法。作为单层生长的已建立细胞系为体外重要过程提供了宝贵的见解,例如增殖,侵袭性,迁移和凋亡1。尽管二维(2D)细胞培养一直是研究哺乳动物细胞对几种环境扰动的反应的传统工具,但推断这些发现以预测组织水平的反应似乎不够令人信服。2D培养的主要局限性在于产生的微环境与乳腺组织本身的微环境有很大不同2。2D培养缺乏细胞与细胞外基质的相互作用,这对于任何组织的生长都至关重要。此外,细胞在单层培养物中经历的拉力会阻碍这些细胞的极性,从而改变细胞信号传导和行为3,4,5。三维(3D)培养系统具有模拟体外体内条件的能力,为癌症研究领域开辟了一条新途径。 在2D细胞培养中丢失的许多关键微环境线索可以使用富含层粘连蛋白的细胞外基质(lrECM)的3D培养物重新建立6。
各种研究已经确定了肿瘤微环境在致癌中的重要性7,8。炎症相关因素是微环境的主要部分。血小板活化因子(PAF)是由各种免疫细胞分泌的磷脂介质,介导多种免疫反应9,10。高水平的PAF由不同的乳腺癌细胞系分泌,并与增殖增强有关11。我们实验室的研究表明,腺泡培养物中PAF的长期存在导致乳腺上皮细胞的转化12。PAF激活PAF受体(PAFR),激活PI3K/Akt信号轴13。据报道,PAFR 也与 EMT、侵袭和转移有关14。
本协议展示了一个模型系统来研究PAF诱导的转化,使用乳腺上皮细胞的3D培养物,如Chakravarty等人之前所描述的那样12。在细胞外基质(3D培养物)上生长的乳腺上皮细胞倾向于形成极化生长停滞球状体。这些被称为腺泡,与乳腺组织的腺泡非常相似,乳腺是乳腺最小的功能单位,在 体内15。 这些球体(图1A,B)由单层紧密堆积的极化上皮细胞组成,围绕空心腔并附着在基底膜上(图1C)。这种形态发生过程已在文献16中得到了很好的描述。当接种在lrECM上时,细胞经历分裂和分化以形成细胞簇,然后从第4天开始极化。到第8天,腺泡由一组与细胞外基质直接接触的极化细胞和封闭在外极化细胞内的一簇非极化细胞组成,与基质没有接触。已知这些未极化的细胞在培养的第12天发生凋亡,形成空心腔。到第16天,形成生长停滞的结构16。
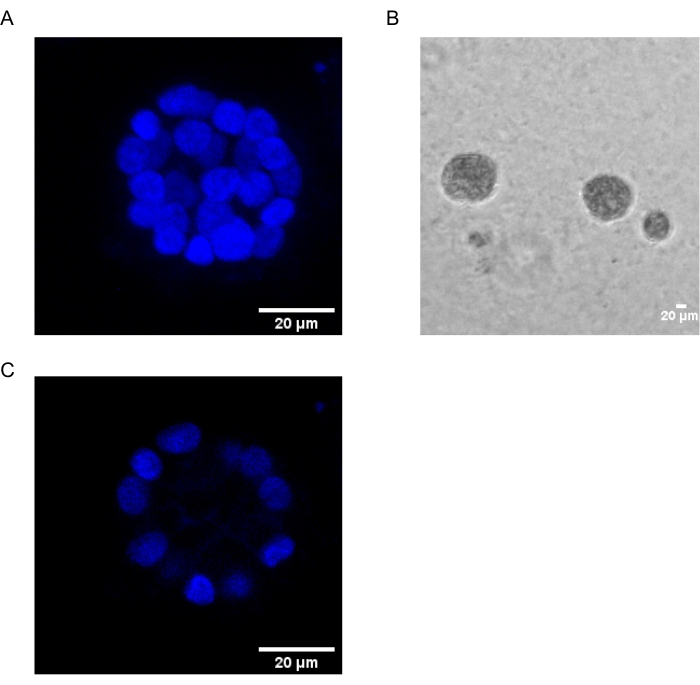
图1:用核染色的腺泡中的细胞核 。 (A)腺泡的3D构造。(B)在基质胶上生长20天的MCF10A腺泡的相衬图像。(C)最中间的部分显示了空心腔的存在。比例尺 = 20 μm。 请点击此处查看此图的大图。
与2D培养不同,腺泡培养有助于通过明显的形态变化区分正常细胞和转化细胞。未转化的乳腺上皮细胞形成具有空腔的腺泡,模仿正常人乳腺腺泡。这些球体在转化时显示出破坏的形态,其特征是极性严重丧失(癌症的标志之一),没有管腔或空腔的破坏(由于细胞凋亡的逃避),这可能是由于各种基因的失调引起的17,18,19,20.这些转化可以使用常用的技术(如免疫荧光)进行研究。因此,3D细胞培养模型可以作为研究乳腺腺泡形态发生和乳腺癌发生的过程的简单方法。建立3D培养系统以了解磷脂介质PAF的作用将有助于高通量临床前药物筛选。
这项工作采用了3D“顶部”培养协议16,21,以研究PAF22诱导的转化。使用免疫荧光研究了腺泡暴露于磷脂介质引起的表型变化。研究中使用了各种极性和上皮到间充质转化(EMT)标记12,16。表1提到了它们的正常定位和转化时的预期表型。
| 抗体 | 标志着 | 正常本地化 | 转化表型 |
| α6-整合素 | 基底外侧 | 基底侧染色较弱 | 强烈的横向/顶端染色 |
| β-连环蛋白 | 细胞-细胞连接 | 基底外侧 | 异常/核或细胞质定位 |
| 维门汀 | EMT | 不存在/弱存在 | 上调 |
表1:研究中使用的标记物。 在存在和不存在PAF治疗的情况下,使用不同的标记物进行定位。
该方法可以最好地用于研究/筛选各种乳腺癌亚型的合理药物和靶基因。这可以提供更接近 体内 情景的药物反应数据,有助于更快、更可靠的药物开发。此外,该系统可用于研究与药物反应和耐药性相关的分子信号传导。
