Una miriade di modelli sono disponibili per studiare la progressione del cancro, ognuno dei quali è unico e rappresenta un sottotipo di questa complessa malattia. Ogni modello fornisce informazioni uniche e preziose sulla biologia del cancro e ha migliorato i mezzi per imitare la condizione reale della malattia. Le linee cellulari consolidate cresciute come monostrato hanno fornito preziose informazioni sui processi vitali in vitro, come proliferazione, invasività, migrazione e apoptosi1. Sebbene la coltura cellulare bidimensionale (2D) sia stata lo strumento tradizionale per studiare la risposta delle cellule di mammifero a diverse perturbazioni ambientali, l’estrapolazione di questi risultati per prevedere le risposte a livello tissutale non sembra sufficientemente convincente. La principale limitazione delle colture 2D è che il microambiente creato differisce in gran parte da quello del tessuto mammario stesso2. La coltura 2D manca dell’interazione delle cellule con la matrice extracellulare, che è vitale per la crescita di qualsiasi tessuto. Inoltre, le forze di trazione sperimentate dalla cellula in colture monostrato ostacolano la polarità di queste cellule, alterando così la segnalazione e il comportamento cellulare 3,4,5. I sistemi di coltura tridimensionali (3D) hanno aperto una nuova strada nel campo della ricerca sul cancro con la loro capacità di imitare le condizioni in vivo in vitro. Molti segnali microambientali cruciali che vengono persi nella coltura cellulare 2D potrebbero essere ristabiliti utilizzando colture 3D di matrice extracellulare ricca di laminina (lrECM)6.
Diversi studi hanno identificato l’importanza del microambiente tumorale nella carcinogenesi 7,8. I fattori associati all’infiammazione sono una parte importante del microambiente. Il fattore attivante piastrinico (PAF) è un mediatore fosfolipidico secreto da varie cellule immunitarie che media le risposte immunitarie multiple 9,10. Alti livelli di PAF sono secreti da diverse linee cellulari di cancro al seno e sono associati a una maggiore proliferazione11. Studi del nostro laboratorio hanno dimostrato che la presenza prolungata di PAF nelle colture acinose porta alla trasformazione delle cellule epiteliali mammarie12. PAF attiva il recettore PAF (PAFR), attivando l’asse di segnalazione PI3K/Akt13. PAFR è anche segnalato per essere associato a EMT, invasione e metastasi14.
Il presente protocollo dimostra un sistema modello per studiare la trasformazione indotta da PAF, utilizzando colture 3D di cellule epiteliali mammarie, come è stato precedentemente descritto da Chakravarty et al.12. Le cellule epiteliali mammarie cresciute sulla matrice extracellulare (colture 3D) tendono a formare sferoidi polarizzati arrestati dalla crescita. Questi sono chiamati acini e assomigliano molto agli acini del tessuto mammario, la più piccola unità funzionale della ghiandola mammaria, in vivo15. Questi sferoidi (Figura 1A,B) sono costituiti da un monostrato di cellule epiteliali polarizzate strettamente imballate che circondano un lume cavo e attaccate alla membrana basale (Figura 1C). Questo processo di morfogenesi è stato ben descritto in letteratura16. Quando seminano su lrECM, le cellule subiscono divisione e differenziazione per formare un gruppo di cellule, che poi si polarizzano dal giorno 4 in poi. Al giorno 8, gli acini sono costituiti da un gruppo di cellule polarizzate che sono in contatto diretto con la matrice extracellulare e un gruppo di cellule non polarizzate racchiuse all’interno delle cellule polarizzate esterne, senza contatto con la matrice. Queste cellule non polarizzate sono note per andare incontro ad apoptosi entro il giorno 12 della coltura, formando un lume cavo. Entro il giorno 16, le strutture in arresto della crescita si formano16.
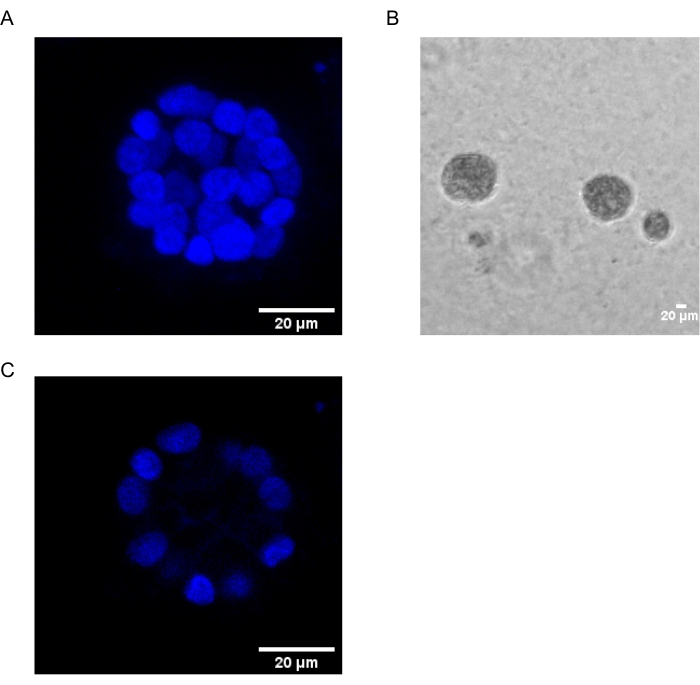
Figura 1: Nuclei di cellule in acini colorati con una colorazione nucleare . (A) Costruzione 3D degli acini. (B) Immagine a contrasto di fase di MCF10A acini coltivati su Matrigel per 20 giorni. (C) La sezione più centrale mostra la presenza di un lume cavo. Barra di scala = 20 μm. Fare clic qui per visualizzare una versione ingrandita di questa figura.
A differenza delle colture 2D, le colture di acinosi aiutano a distinguere le cellule normali e trasformate attraverso apparenti cambiamenti morfologici. Le cellule epiteliali mammarie non trasformate formano acini con un lume cavo, imitando i normali acini del seno umano. Questi sferoidi, dopo la trasformazione, mostrano una morfologia perturbata caratterizzata da una grave perdita di polarità (uno dei segni distintivi del cancro), assenza di un lume o rottura del lume cavo (a causa dell’evasione dell’apoptosi) che può essere indotta a causa della deregolazione di vari geni17,18,19,20 . Queste trasformazioni possono essere studiate utilizzando tecniche comunemente usate come l’immunofluorescenza. Pertanto, il modello di coltura cellulare 3D può funzionare come un metodo semplice per studiare il processo di morfogenesi acinogenesi del seno e la carcinogenesi del seno. Stabilire un sistema di coltura 3D per comprendere l’effetto di un mediatore dei fosfolipidi, PAF, aiuterà nello screening preclinico dei farmaci ad alto rendimento.
Questo lavoro ha adattato il protocollo di coltura 3D “on top”16,21 per studiare la trasformazione indotta da PAF 22. I cambiamenti fenotipici indotti dall’esposizione degli acini al mediatore fosfolipidico sono stati studiati utilizzando l’immunofluorescenza. Nello studio sono stati utilizzati vari marcatori di polarità ed epiteliale-transizione mesenchimale (EMT)12,16. La Tabella 1 menziona la loro normale localizzazione e il loro fenotipo atteso dopo la trasformazione.
| Anticorpi | Marchi | Localizzazione normale | Fenotipo trasformato |
| α6-integrina | Basolaterale | Basali con debole macchia laterale | Forte macchia laterale / apicale |
| β-catenina | Giunzione cellula-cellula | Basolaterale | Localizzazione anomala / nucleare o citoplasmatica |
| Vimentino | Emt | Presenza assente / debole | Up-regulation |
Tabella 1: Marcatori utilizzati nello studio. Diversi marcatori utilizzati con la loro localizzazione in presenza e assenza di trattamento PAF.
Questo metodo può essere utilizzato al meglio per studiare / selezionare farmaci plausibili e geni bersaglio per vari sottotipi di cancro al seno. Ciò può fornire dati di risposta ai farmaci più vicini allo scenario in vivo , contribuendo a uno sviluppo più rapido e affidabile del farmaco. Inoltre, questo sistema può essere utilizzato per studiare la segnalazione molecolare associata alla risposta ai farmaci e alla resistenza ai farmaci.
