2011 में, शोधकर्ताओं ने एक कम्प्यूटेशनल विधि का प्रस्ताव दिया और डिजिटल सिंथेटिक जीन सर्किट1 के स्वचालित डिजाइन के लिए सॉफ्टवेयर का एक संबंधित टुकड़ा विकसित किया। एक उपयोगकर्ता को इनपुट (तीन या चार) की संख्या निर्दिष्ट करनी थी और सर्किट सत्य तालिका भरनी थी; इसने इलेक्ट्रॉनिक्स से तकनीकों का उपयोग करके सर्किट संरचना प्राप्त करने के लिए सभी आवश्यक जानकारी प्रदान की। सत्य तालिका का कर्णौघ मानचित्र विधि2 के माध्यम से दो बूलियन सूत्रों में अनुवाद किया गया था। प्रत्येक बूलियन सूत्र खंडों से बना होता है जो सर्किट इनपुट और उनके निषेधों (शाब्दिक) के बीच तर्क संचालन (योग या गुणन) का वर्णन करते हैं। सर्किट आउटपुट की गणना करने के लिए खंड, उनकी बारी में, या तो अभिव्यक्त (ओआर) या गुणा (एएनडी) होते हैं। प्रत्येक परिपथ को इसके दो संगत सूत्रों में से किसी एक के अनुसार महसूस किया जा सकता है: एक पीओएस (रकम का उत्पाद) रूप में लिखा गया है और दूसरा एसओपी (उत्पादों का योग) प्रतिनिधित्व में लिखा गया है। पूर्व में खंडों (यानी, बूलियन गेट) का गुणन होता है जिसमें शाब्दिक का तर्क योग होता है। उत्तरार्द्ध, इसके विपरीत, खंडों का एक योग है जहां शाब्दिक गुणा किए जाते हैं।
इलेक्ट्रिक सर्किट को ब्रेडबोर्ड पर, शारीरिक रूप से विभिन्न द्वारों को एक साथ वायर करके महसूस किया जा सकता है। विद्युत प्रवाह फाटकों के बीच संकेतों के आदान-प्रदान की अनुमति देता है, जिससे आउटपुट की गणना होती है।
जीव विज्ञान में, स्थिति अधिक जटिल है। एक बूलियन गेट को प्रतिलेखन इकाई (टीयू; यानी, यूकेरियोटिक कोशिकाओं के अंदर अनुक्रम “प्रमोटर-कोडिंग क्षेत्र-टर्मिनेटर” के रूप में महसूस किया जा सकता है, जहां प्रतिलेखन या अनुवाद (या दोनों) को विनियमित किया जाता है। इस प्रकार, कम से कम दो प्रकार के अणु एक जैविक वायरिंग स्थापित करते हैं: प्रतिलेखन कारक प्रोटीन और गैर-कोडिंग, एंटीसेंस आरएनए1।
एक जीन डिजिटल सर्किट को गेट की दो या तीन परतों में व्यवस्थित किया जाता है, अर्थात्: 1) इनपुट परत, जो हां (बफर) से बनी होती है और गेट नहीं होती है और इनपुट रसायनों को वायरिंग अणुओं में परिवर्तित करती है; 2) आंतरिक परत, जिसमें उतने ही टीयू होते हैं जितने कि संबंधित बूलियन सूत्र में खंड होते हैं। यदि सर्किट एसओपी सूत्र के अनुसार डिज़ाइन किया गया है, तो आंतरिक परत में प्रत्येक खंड तथाकथित वितरित आउटपुट आर्किटेक्चर में सर्किट आउटपुट (जैसे, प्रतिदीप्ति) का उत्पादन करेगा। यदि योग (पीओएस) सूत्र के उत्पाद का उपयोग किया जाता है, तो एक 3) अंतिम परत की आवश्यकता होती है, जिसमें आंतरिक परत से तारों के अणुओं को इकट्ठा करने वाला एक एकल गुणक द्वार होगा।
कुल मिलाकर, सिंथेटिक जीव विज्ञान में, एक ही सर्किट के लिए कई अलग-अलग योजनाएं तैयार की जा सकती हैं। वे टीयू और वायरिंग अणुओं दोनों की संख्या और प्रकार में भिन्न होते हैं। खमीर कोशिकाओं में लागू किए जाने वाले सबसे आसान समाधान को चुनने के लिए, प्रत्येक सर्किट डिज़ाइन एक जटिलता स्कोर एस के साथ जुड़ा हुआ है, जिसे
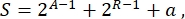
जहां ए सक्रियकर्ताओं की संख्या का प्रतिनिधित्व करता है, आर दमनकारियों की संख्या का प्रतिनिधित्व करता है, और ए एंटीसेंस आरएनए अणुओं की मात्रा है। यदि या तो सक्रियकर्ता या दमनकारी सर्किट से अनुपस्थित हैं, तो एस में उनका योगदान शून्य है। इसलिए, प्रयोगशाला (उच्च एस) में एक सर्किट योजना का एहसास करना अधिक कठिन है जब इसे उच्च संख्या में ऑर्थोगोनल प्रतिलेखन कारकों की आवश्यकता होती है। इसका मतलब है कि डिजिटल सर्किट के अंदर पूर्ण तारों का एहसास करने के लिए नए सक्रियकर्ताओं और दमनकारियों को नए सिरे से इंजीनियर किया जाएगा। सिद्धांत रूप में, नोवेल डीएनए-बाइंडिंग प्रोटीन को जिंक फिंगर प्रोटीन3 और टीएएल प्रभावकों4 का उपयोग करके टेम्पलेट के रूप में इकट्ठा किया जा सकता है। हालांकि, यह विकल्प बहुत कठिन और समय लेने वाला प्रतीत होता है; इसलिए, जटिल जीन सर्किट को अंतिम रूप देने के लिए ज्यादातर छोटे आरएनए और अनुवाद विनियमन पर भरोसा करना चाहिए।
मूल रूप से, यह विधि बैक्टीरिया में डिजिटल सर्किट बनाने के लिए विकसित की गई थी। दरअसल, यूकेरियोटिक कोशिकाओं में, एंटीसेंस आरएनए के बजाय, माइक्रोआरएनए (एमआईआरएनए) या छोटे हस्तक्षेप करने वाले आरएनए (एसआईआरएनए) 5 के बारे में बात करना अधिक उपयुक्त है। हालांकि, आरएनएआई मार्ग खमीर एस सेरेविसिया में मौजूद नहीं है। इसलिए, किसी को पूरी तरह से ट्रांसक्रिप्शनल नेटवर्क का विकल्प चुनना चाहिए। मान लीजिए कि एक सर्किट को पांच सक्रियकर्ताओं और पांच दमनकारियों की आवश्यकता होती है; इसका जटिलता स्कोर एस = 32 होगा। सर्किट जटिलता को 10 प्रतिलेखन कारकों को एक सक्रियण डोमेन (एडी) से जुड़े एकल डीसीएएस 96 (न्यूक्लियस की कमी वाले कैस 9) के साथ बदलकर कम किया जा सकता है। जैसा कि7 में दिखाया गया है, डीसीएएस 9-एडी टाटा बॉक्स और टीएसएस (ट्रांसक्रिप्शन स्टार्ट साइट) के बीच प्रमोटर को बांधते समय खमीर में एक दमनकारी के रूप में काम करता है और टाटा बॉक्स के ऊपर अच्छी तरह से बांधने पर एक उत्प्रेरक के रूप में काम करता है। इस प्रकार, एस = 11 के कुल जटिलता स्कोर के लिए 10 प्रतिलेखन कारकों को एकल डीसीएएस 9-एडी संलयन प्रोटीन और 10 एसजीआरएनए (एकल गाइड आरएनए) के साथ प्रतिस्थापित किया जा सकता है। दस एसजीआरएनए को संश्लेषित करना त्वरित और आसान है, जबकि, जैसा कि पहले टिप्पणी की गई थी, 10 प्रोटीन ों की असेंबली बहुत लंबे और अधिक जटिल काम की मांग करेगी।
वैकल्पिक रूप से, कोई दो ऑर्थोगोनल डीसीएएस प्रोटीन (जैसे, डीसीएएस 9 और डीसीएएस 12 ए) का उपयोग कर सकता है: एक एडी में फ्यूज करने के लिए, और दूसरा नंगे या दमन डोमेन के साथ संयोजन में। जटिलता स्कोर केवल एक इकाई (एस = 12) से बढ़ेगा। इसलिए, CRISPR-dCas सिस्टम S. Cerevisiae में बहुत जटिल जीन डिजिटल सर्किट के निर्माण की कुंजी हैं।
यह पेपर खमीर में dCas9- और dCas12a-आधारित रिप्रेसर्स और एक्टिवेटर्स दोनों की दक्षता को गहराई से दर्शाता है। परिणाम बताते हैं कि वे अपनी गतिविधि को अनुकूलित करने के लिए एसजीआरएनए की उच्च मात्रा की मांग नहीं करते हैं, इसलिए एपिसोमल प्लास्मिड को अधिमानतः टाला जाता है। इसके अलावा, डीसीएएस 9-आधारित सक्रियकर्ता एक मचान आरएनए (एससीआरएनए) का उपयोग करते समय कहीं अधिक प्रभावी होते हैं जो वीपी 64 एडी की प्रतियां भर्ती करता है। इसके विपरीत, मजबूत वीपीआर एडी से सीधे जुड़े होने पर डीसीएएस 12 ए अच्छी तरह से काम करता है। इसके अलावा, एक सिंथेटिक सक्रिय प्रमोटर एक्टिवेटर के कॉन्फ़िगरेशन के आधार पर लक्ष्य साइटों की एक चर संख्या की मांग करता है (उदाहरण के लिए, डीसीएएस 12 ए-वीपीआर का उपयोग करते समय तीन, डीसीएएस 9-वीपी 64 के लिए छह, और डीसीएएस 9 और एससीआरएनए के साथ केवल एक)। एक दमनकारी के रूप में, प्रमोटर के बजाय कोडिंग क्षेत्र को बांधते समय डीसीएएस 12 ए अधिक तीक्ष्ण दिखाई देता है।
एक दोष के रूप में, हालांकि, CRISPR-dCas9 / dCas12a सीधे रसायनों के साथ बातचीत नहीं करते हैं। इसलिए, इनपुट परत में उनका कोई उपयोग नहीं हो सकता है। इस कारण से, एंटी-सीआरआईएसपीआर प्रोटीन (एसीआर) युक्त वैकल्पिक बूलियन गेट डिजाइनों की जांच की गई है। एसीआरएस (डी) सीएएस प्रोटीन पर कार्य करते हैं औरउनके काम को रोकते हैं। इसलिए, वे CRISPR-(d) CAS सिस्टम की गतिविधि को संशोधित करने का एक साधन हैं। यह पेपर टाइप II Acrs और (d) Cas9 के बीच बातचीत का पूरी तरह से विश्लेषण करता है, साथ ही साथ टाइप V Acrs और (d) Cas12a में S. cerevisiae में। चूंकि एसीआरएस सीएएस प्रोटीन की तुलना में बहुत छोटे होते हैं, इसलिए एस्ट्रोजेन β-एस्ट्राडियोल के प्रति उत्तरदायी एक नॉट गेट मानव एस्ट्रोजन रिसेप्टर 9-एचबीडी (एचईआर) -से एसीआरआईआईए 4 के हार्मोन-बाइंडिंग डोमेन को फ्यूज करके बनाया गया था। इसके अलावा, गैलेक्टोज के साथ प्रेरण पर डीसीएएस 12 ए (-एडी) और एसीआरवीए को संवैधानिक रूप से व्यक्त करने वाले मुट्ठी भर हां और नॉट गेट्स महसूस किए गए थे। वर्तमान में, ये द्वार केवल अवधारणा के प्रमाण के रूप में कार्य करते हैं। हालांकि, वे खमीर कोशिकाओं में सिंथेटिक जीन डिजिटल सर्किट के कम्प्यूटेशनल स्वचालित डिजाइन को पूरा करने के लिए एल्गोरिदम के गहरे पुनर्विचार की दिशा में पहले कदम का भी प्रतिनिधित्व करते हैं।
