If the protocol has been successfully completed (Figure 1), the sample will appear clear and flat after heat denaturation; any folding or wrinkling indicates incomplete homogenization. A successfully expanded sample will be 3-4.5-fold larger than before expansion in 1x PBS and 8-11-fold larger when fully expanded in ddH2O. Figure 3 shows example pre- and post-expansion images of 5 µm thick FFPE human kidney sample processed using this protocol and successfully expanded over 8-fold. The tissue was first stained with antibodies for ACTN4 along with DAPI to visualize the nuclear DNA. The specimen was then imaged using a spinning disk confocal microscope (Figure 3A). The tissue was treated following the above protocol, including the post-expansion staining of the same targets, and fully expanded in water (Figure 3B) before re-imaging. Following heat denaturation, biomolecules other than proteins may be stained for and imaged as well, as shown in Figure 4. Lipophilic dyes can be applied to reveal the cell and mitochondria membrane structures in the mouse brain (Figure 4A). Additionally, nucleic acids may be imaged through FISH, as in the FFPE normal human lymph node tissue shown in Figure 4B,C.
Figure 5 demonstrates a likely outcome if the sample is not properly homogenized. Following expansion, the tissue was stained with antibodies for ACTN4 and vimentin, along with DAPI to visualize the nuclear DNA and wheat germ agglutinin (WGA) to label the carbohydrates. The tissue was expanded 3.5-fold in PBS before imaging using a spinning disk confocal microscope. In one kidney section (Figure 5A,C), the crack-free expansion of a glomerulus can be seen. In a separate section (Figure 5B,D), cracking can be clearly seen in the tissue.
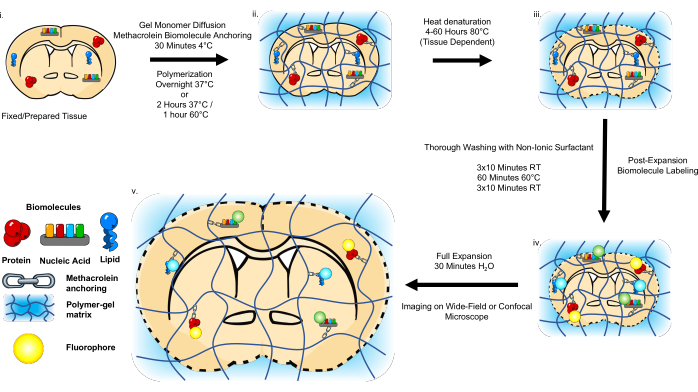
Figure 1: Schematic of the Magnify workflow. The pre-processing of clinically archived tissue slides is first performed based on the storage format. Free-floating paraformaldehyde-fixed sections need only be washed in PBS. The samples are then incubated in a gel monomer solution, including methacrolein to anchor the biomolecules to the hydrogel. In situ polymerization is performed prior to heat denaturation with urea, SDS, and EDTA. The samples are then thoroughly washed and stained using conventional immunostaining protocols, FISH protocols, or lipophilic dyes. The samples are then expanded in pure water before imaging. This figure has been modified from Klimas et al.17. Please click here to view a larger version of this figure.
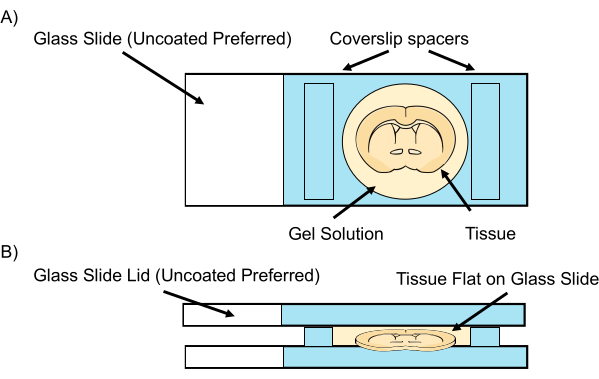
Figure 2: Tissue sample gelation chamber. (A) On each side of the tissue, two spacers, such as two pieces of #1.0 cover glass, are placed before the gel monomer solution is allowed to diffuse at 4 °C. To prevent compression, the spacers should be thicker than the tissue slices. (B) A lid, such as a second glass slide, is used to cover the sample before polymerization at 37 °C. Please click here to view a larger version of this figure.
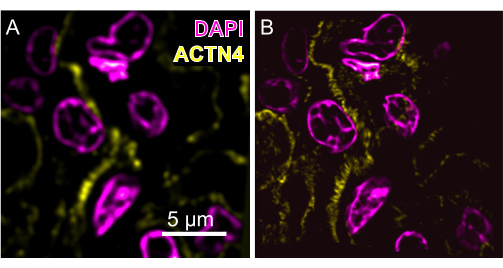
Figure 3: Example pre-expansion images of a human kidney tissue section. (A) An image taken at 60x magnification (1.4 NA) compared to (B) an image of the same field of view post-expansion with Magnify taken at 40x magnification (1.15 NA, expansion factor: 8.15x). Magenta, DAPI; Yellow, ACTN4. The maximum intensity of the post-expansion images is projected over 25 frames. Please click here to view a larger version of this figure.
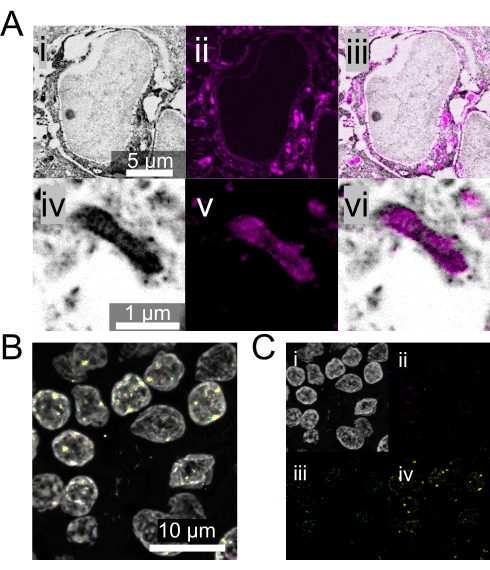
Figure 4: Alternative biomolecule staining strategies with Magnify. (A) (i and iv) NHS-ester pan-protein labeling of fully-expanded mouse brain tissue. (ii and v) Lipophilic dye (DiD) labeling of the same mouse brain tissue. (iii and vi) The channels merged. (B) DNA FISH with Magnify using FFPE normal human lymph node tissue. Expansion factor: 3.5x in 1x PBS. White, DAPI; Magenta, serine/threonine kinase 1 gene; Blue, APC/C activator protein CDH1 gene; Yellow, human satellite sequence S4. (C) Individual channels associated with B. All the images were obtained using a spinning disk confocal microscope at 40x magnification (1.15 NA). Please click here to view a larger version of this figure.
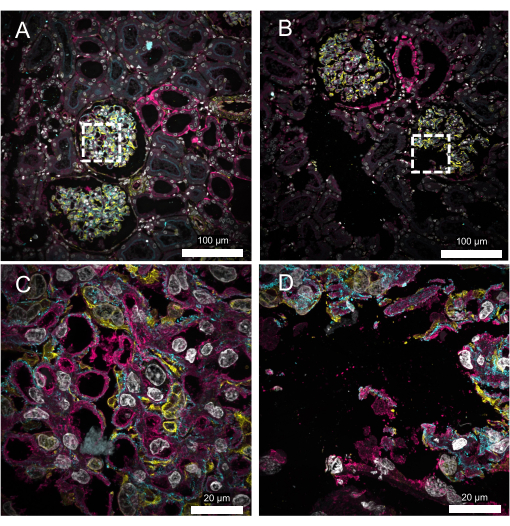
Figure 5: Representative results of 5 µm thick FFPE kidney samples. (A) Crack-free expansion. White, DAPI; Yellow, vimentin; Cyan, alpha-actinin 4; Magenta, wheat germ agglutinin. (B) Expanded kidney section exhibiting cracking. Cracking, distortions, and the loss of labeled targets can be the result of inadequate anchoring and/or homogenization. (C,D) Zoomed-in images of the boxed regions in A and B, respectively. All the images were obtained using a spinning disk confocal microscope at 10x (A,B; 0.45 NA) or 60x (C,D; 1.2 NA) magnification. Please click here to view a larger version of this figure.
Table 1: Gel monomer solution composition. Please click here to download this Table.
Table 2: Homogenization buffer composition. Please click here to download this Table.
Table 3: Summary of the methacrolein and homogenization conditions for validated tissues. Please click here to download this Table.
