Monitoring of the fish after cryoinjury
To determine the effect of myomere cryoinjury on animals, a video recording of control and cryoinjured fish at 1 days post-cryoinjury (dpci) and 5 dpci was performed. Each group contained five fish. At 1 dpci, the cryoinjured fish were swimming less actively, but they did not display any abnormal movements, such as swirling, convolution, or reduced equilibrium (Video 1). In the husbandry system, their position in the tank and food intake were similar to those of the uninjured fish. Normal behavior persisted throughout the following days, as exemplified by the video at 5 dpci (Video 2). In conclusion, the cryoinjury procedure of the caudal peduncle did not severely affect the well-being of the animals.
Histological analysis of the caudal peduncle sections
To assess the extent of the injury, the time point of 4 dpci was selected, as this is when the myofiber debris has been completely resorbed in the wound. To analyze the effects of cryoinjury along the dorso-ventral and anterior-posterior axes of the body, two groups of fish were used (i.e., coronal and transversal sections of the caudal peduncle, respectively) (Figure 1F).
The sections were analyzed by trichrome staining composed of Aniline Blue, Acid Fuchsin, and Orange G (AFOG). Using this combination of reagents, intact muscles were shown in orange, the spinal cord in dark red, and the collagenous matrix in blue. To determine the number of damaged myomeres, which are the metameric units of the fish musculature, a series of sections were analyzed (Figure 2). Myomere boundaries, called myocommata, were identified by collagen deposition, as detected by blue coloration. The damaged areas were determined by the absence of orange staining. A closer examination of specimens with evident myocommata revealed that approximately four consecutive myomeres were damaged, as inferred from the lack of orange staining (n, number of fish = 4; Figure 3A,A'). The uninjured side of the same fish served as the internal reference.
To examine the depth of the wound perpendicular to the body axis, transversal sections were prepared using zebrafish at 4 dpci and 7 dpci. The latter time point corresponds to the activation of the myogenic program and, thus, the onset of muscle regeneration. AFOG staining of these specimens displayed an extensive lack of orange staining in the cryoinjured flank of the body, demarcating the zone of degenerated skeletal muscle (Figure 3B,C). At 4 dpci and 7 dpci, the wound area spanned tissues from the skin toward the vertical septum. This demonstrates that the cryoinjury method deeply targeted one lateral half of the caudal peduncle, which remained devoid of functional muscle for 7 days after the procedure. Taken together, four myomeres were profoundly damaged on one side of the caudal peduncle.
Immunofluorescence analysis of the transversal sections
To assess the dynamics of muscle regeneration, experimental groups of fish were euthanized at 4 dpci, 7 dpci, 10 dpci, and 30 dpci. The transversal sections of the caudal peduncle were labeled by multicolor fluorescence staining using Phalloidin (which binds to filamentous actin [F-actin]), Tropomyosin-1 antibody, which detects a sarcomere protein, and DAPI, which labels nuclei. At all the time points, the uninjured half of the body provided an internal control; both F-actin and Tropomyosin 1 were strongly detected in the uninjured control parts, indicating undamaged tissue (Figure 4).
At 4 dpci, the injured side of the caudal peduncle contained abundant DAPI-positive cells, but little to no F-actin and Tropomyosin 1 immunofluorescence was observed, indicating the wound zone with degenerated muscles (Figure 4A–B'). At 7 dpci, Tropomyosin 1 and F-actin could be detected in a part of the wound close to the vertical body midline (Figure 4C–D'). This expression pattern demarcates the position where the formation of new myofibers begins in the caudal peduncle. At 10 dpci, both muscle markers expanded toward the surface of the body, suggesting progressing regeneration of the skeletal muscle (Figure 4E–F'). At 30 dpci, both sides of the body displayed a similar distribution of F-actin staining (Figure 4G–H'). This finding indicates that the skeletal muscle was efficiently restored after the cryoinjury of the caudal peduncle.
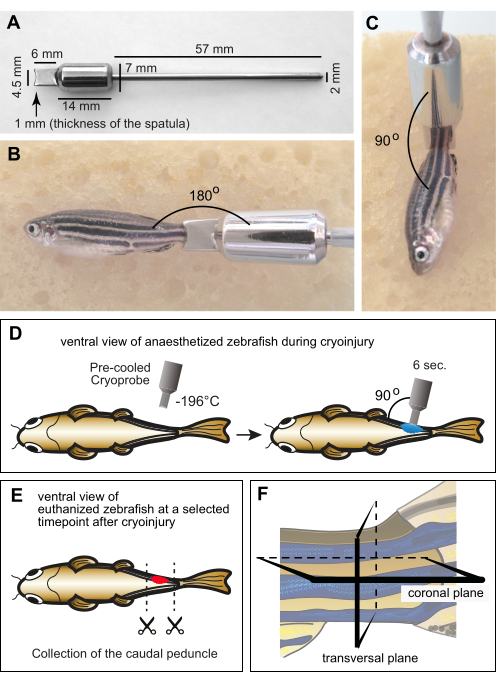
Figure 1: Experimental setup for myomere cryoinjury. (A) Dimensions of the custom-manufactured cryoprobe made of stainless steel. The distal part of the instrument consists of a spatula with a concave edge at the depth of 1 mm to account for the curvature of the zebrafish body. The middle part of the tool comprises a cylinder that functions as a weight and a reservoir to maintain the low temperature of the spatula during the procedure. The proximal end of the instrument is in the form of a thin metal handle. (B,C) Anesthetized adult fish on a moist sponge with the cryoprobe on the caudal peduncle. The probe was at room temperature. (B) The margin of the probe is placed horizontally in the vicinity of the caudal peduncle to display the relative size between the fish and the tool. (C) For cryoinjury, the tip of the tool is positioned perpendicular to the fish. (D) Schematic illustration of the cryoinjury procedure from the ventral side of the fish to show the manipulations in a comprehensive manner. The cryoprobe was precooled in liquid nitrogen and immediately placed on one side of the fish for 6 s. (E) At a specific time point after cryoinjury, the fish were euthanized, and their caudal peduncles were collected for fixation. (F) The fixed material was histologically processed and sectioned along the coronal or transversal planes. Please click here to view a larger version of this figure.
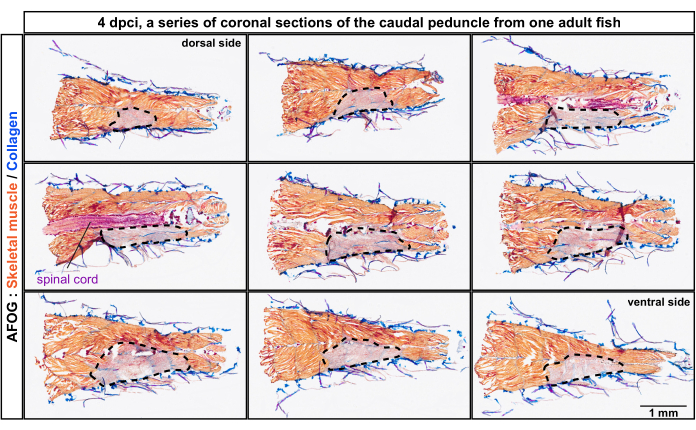
Figure 2: Histological analysis of damaged myomeres in the caudal peduncle from the dorsal to the ventral position of the body. AFOG staining of a series of coronal sections at 4 days post-cryoinjury (dpci). The sections are from the dorsal toward the ventral side, as indicated on the top of the first and the last panel. The sections are non-adjacent, with an interval of approximately 150 µm between them. The uninjured muscle is detected by orange staining of the muscle, whereas the injured tissue lacks this staining and appears grayish (area encircled with a dashed line). Collagen-containing tissues, such as the skin, are stained in blue. The spinal cord appears as a rod-like structure and is stained in red. Number of fish, n = 4. Please click here to view a larger version of this figure.
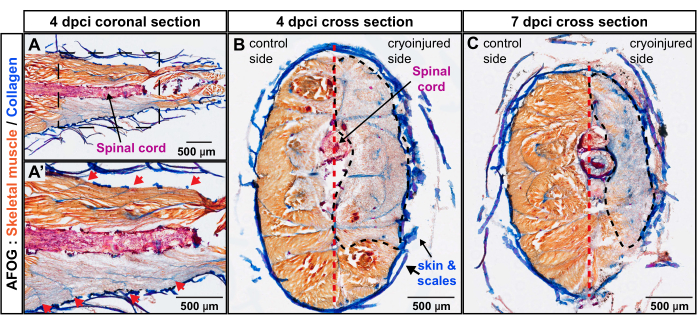
Figure 3: Assessment of injury depth in the caudal peduncle using AFOG staining. (A,A') The coronal section is at the level of the spinal cord (a red-stained horizontal rod). The bottom image shows a magnified area encompassed with a frame in the upper image. The sequential myomere boundaries appear as collagenous stripes (blue) positioned obliquely to the spinal cord (red arrows in the magnified image A').(B,C) The cross-sections display the uninjured flank with orange-stained muscles and the cryoinjured flank with grayish staining. The damaged area is encircled with a black dashed line. The vertical septum (depicted with a red dashed line) subdivides the body into the control and cryoinjured sides. Number of fish, n = 4 per time point. Please click here to view a larger version of this figure.
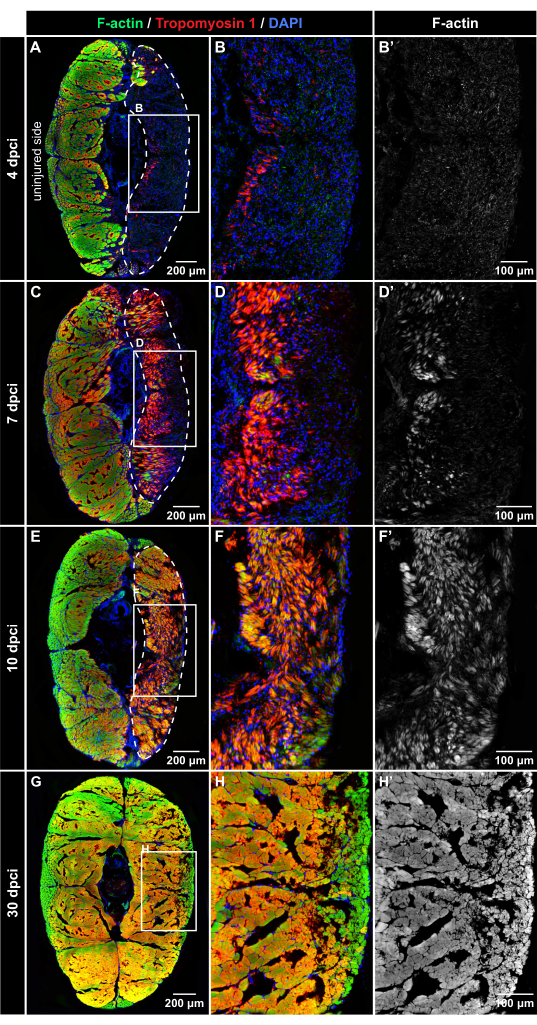
Figure 4: Immunofluorescent detection of muscle proteins after cryoinjury. Fluorescence staining of cross-sections at 4 dpci, 7 dpci, 10 dpci, and 30 dpci, as labeled on the left side and the top of the panels (A–B'). At 4 dpci, the injured tissue (encircled with the dashed line) is DAPI-positive (blue) but devoid of Phalloidin staining (green) or Tropomyosin-1 immunoreactivity (red), suggesting degeneration of the muscle fibers after cryoinjury. (C–D') At 7 dpci, both muscle markers progressively emerge in the wounded area, indicating the regenerative process. Tropomyosin-1 appears more intense than F-actin in the newly formed fibers. (E–F') At 10 dpci, the injury zone is filled with new myofibers that display a higher intensity of Tropomyosin-1 immunoreactivity in comparison to F-actin. (G–H') At 30 dpci, a similar pattern of myofibers is detected in both sides of the body. The frames in panels A, C, E, and H encompass the areas that are magnified in the adjacent images to the right. Dermal scales, emanating fluorescence outside of the myomere, were erased from the images using Adobe Photoshop. Number of fish, n = 4 per time point. Please click here to view a larger version of this figure.
