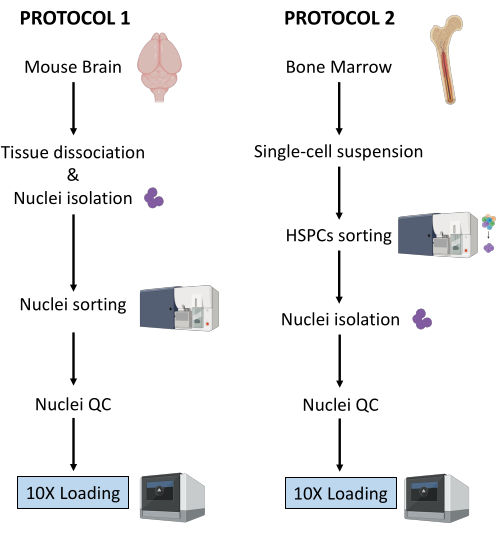
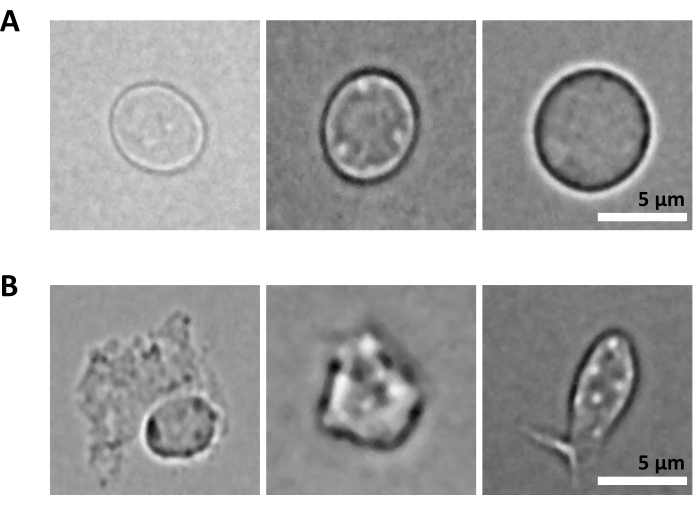
Försöksmetoderna utfördes i enlighet med de protokoll som godkänts av kommittén för djurförsöksetik (CETEA). För isolering av hjärnkärnor användes 3 månader gamla C57BL/6-möss. För benmärgsisoleringen användes 8 veckor gamla C57BL/6J-möss av honkön som vägde 18 g. 1. Rening av kärnor från mushjärna OBS: Använd latex- eller nitrilhandskar hela tiden under proceduren. Det rekommenderas starkt att ha två personer som utför experimentet, att låta steg 1 till 3 (dvs. beredning av suspensionen av enstaka kärnor) utföras av en person, och steg 4 (dvs. förberedelse av sorteraren) utförs parallellt av en annan person. Eftersom protokollet är mycket tidskänsligt är det viktigt att minimera provbearbetningstiden genom att ha sorteraren redo så snart suspensionen med enstaka kärnor har förberetts. Beredning av reagenser och materialSterilisera dissektionsverktygen försiktigt i autoklav (vid 121 °C i 20 minuter) och tvätta dem med etanol 70 % strax före användning. Förbered en petriskål per prov, fylld med 2-3 ml iskall 1x Dulbeccos fosfatbuffrade saltlösning (DPBS). Kyl mikrocentrifugen till 4 °C, fyll en hink med is och lägg glashomogenisatorn på is. Förbered nuclei lysis buffer genom att tillsätta digitonin för en slutlig koncentration på 0,01 %, 10 ml per sample. Bered färgningsbufferten genom att tillsätta RNashämmare till cellfärgningsbufferten för en slutlig koncentration på 0,2 E/μL, 20 ml per prov. Bered DPBS 0,04 % BSA genom att tillsätta RNashämmare för en slutlig koncentration på 0,2 E/μL, 2 ml per prov. Bered 1 ml utspädd kärnbuffert enligt Multiome-protokollet25. Förvara alla reagenser och prover på is. Dissektion av vävnadOffra möss med hjälp av protokoll som godkänts av institutionen. I detta protokoll halshöggs mössen efter en överdos av ketamin/xylazin. Klipp av mushuvudet med en sax och ta bort hjärnan från skallen enligt beskrivningen i Meyerhoff et al.26. Överför omedelbart hjärnan till en petriskål tillagad med den iskalla 1x DPBS under ett lysdiodbelyst (LED)-belyst stereomikroskop. Skär hjärnvävnaden med en skalpell för att separera hjärnområden av intresse (t.ex. Entorhinal cortex, hippocampus, prefrontal cortex) och överför varje region till en separat petriskål som innehåller iskall 1x DPBS. Håll dig på isen. Finhacka vävnaden i <0,5 cm stora bitar med en skalpell för att underlätta homogeniseringen i nästa steg. Med en P1000 mikropipett överför du den malda vävnaden och 1x DPBS från petriskålen till ett 1,5 ml rör. Se till att använda tuber av plast med låg proteinbindningsgrad. Låt vävnadsbitarna separeras av gravitationen. Ta försiktigt bort överskottet av 1x DPBS med en P1000 mikropipett.OBS: Efter detta steg är det möjligt att snäppfrysa den malda vävnaden genom att överföra proteinets lågbindande rör till torris och sedan förvara vid -80 °C tills kärnisoleringen fortsätter. Isolering av kärnorFyll glaset med 2 ml iskall nuclei lysis buffer med 0,01 % digitonin. Tillsätt vävnadsbitarna i dounce.OBS: om du arbetar med färskfryst vävnad, tillsätt den malda frysta vävnaden direkt till kärnlysbufferten 0.01 % digitonin; Låt inte näsduken tina innan. Homogenisera med hjälp av en homogenisator av glasduk 25 gånger med mortelstöt A och sedan 25 gånger med mortelstöt B. Överför homogenatet till ett 15 ml rör. Tillsätt ytterligare 2 ml iskall nuclei lysis buffer med 0,01 % digitonin och inkubera på is i 5 min. Centrifugera kärnorna vid 500 x g i 5 minuter vid 4 °C. Ta bort supernatanten med en mikropipett och tillsätt 4 ml iskall nuclei lysis buffer med 0,01 % digitonin. Inkubera på is i 5 minuter och filtrera genom en 40 μm cellsil. Centrifugera kärnorna vid 500 x g i 5 minuter vid 4 °C och avlägsna supernatanten med en mikropipett. Tillsätt 4 ml färgningsbuffert för att tvätta kärnorna och centrifugera vid 500 x g i 5 minuter vid 4 °C. Ta bort supernatanten med en mikropipett och återsuspendera pelleten i 4 ml färgningsbuffert. Filtrera genom en 40 μm cellsil och centrifugera vid 500 x g i 5 minuter vid 4 °C. Återsuspendera i 1 ml PBS med 0,04 % BSA. Räkna kärnor för att säkerställa konsistens i vävnads-/kärnberedningen över olika prover. Den förväntas få liknande antal kärnor från samma hjärnregioner:Tillsätt 10 μL 0,4 % trypanblå till en tom 0,5 ml tub. Tillsätt 10 μl av kärnorna och blanda 5 gånger genom pipettering. Räkna kärnor med hjälp av en automatiserad cellräknare enligt leverantörens rekommendationer. Håll kärnorna på is. Förbered kärnor för sortering.OBS: De extraherade kärnorna innehåller 7-AAD, och denna färgning används för rening av fluorescensaktiverad cellsorterare (FACS).Överför 100 μL kärnor till ett FACS-rör för ofärgad kontroll. Tillsätt 10 μl 7-AAD till de återstående kärnorna och håll 5 minuter vid 4 °C. Sortera minst 0,5 x 106 kärnor efter FACS för att eliminera dubbletter och skräp. Sortering av kärnor med hjälp av en FACSOBS: Kärnsortering kan utföras på en mängd olika cellsorterare, men proceduren för att använda instrumenten BD FACSAria Fusion eller BD FACSAria III beskrivs här. Det rekommenderas starkt att kalibreringen och installationen av cellsorteraren utförs under överinseende eller av en erfaren användare av instrumentet. För att minska bearbetningstiden för proverna är det viktigt att ha sorteraren redo så snart suspensionen med enstaka kärnor har beretts.Kalibrering av FACS-instrumentSlå på cellsorteraren och datorn. När programvaran är ansluten till instrumentet, starta den fluidiska startproceduren. Välj Cytometer > vätskestart i huvudmenyn och följ de fyra stegen. Klicka på Klar när du har slutfört varje. Sätt i 70 μm-munstycket, slå på strömmen och låt strömmen stabiliseras i 15 minuter. Justera amplituden för att få droppbildning och klicka på Sweet Spot. Sätt i neutraldensitetsfiltret (ND) 1.0 och öppna gränssnittet för cytometerinställning och spårning (CST). Daglig kvalitetskontroll: Späd ut CST-pärlor i FACS-medium (se leverantörens rekommendationer) och utför CST-kontroll. När du är klar, byt ut ND 1.0 mot ND 2.0. Späd ut accudrops i FACS-medium (se leverantörens rekommendationer) och utför droppfördröjning enligt beskrivningen i steg 6 till 10. I experimentmallen väljer du experimentet Accudrop Drop Delay och öppnar sorteringslayouten för röret. Inuti det nedre kamerafönstret klickar du på Voltage och sedan på Optiskt filter för att göra det möjligt att applicera laddning på avböjningsplattorna och använda ett specifikt optiskt filter framför kameran. Se till att kvadranten på höger sida visar 100. Justera vid behov den röda laserskruven för att optimera lasereffekten. Justera flödeshastigheten för att nå en hastighet på 1 000 till 3 000 händelser per sekund. Klicka på Sortera och avbryt. Se till att den vänstra kvadranten är lika med 100 och att den högra kvadranten är 0. Om den vänstra kvadranten är under 95, utför Auto Delay. Klicka på Spänning och sedan på Testsortering. Kontrollera kvaliteten på sidoströmmarna som deponeras i uppsamlingsrören. Om det behövs, justera sidoströmmarnas position genom att flytta reglagen. Uppsättning av FACS-instrument för sortering av kärnor.Börja samla in de ofärgade kärnorna. Dessa används för att definiera de främre och sidospridningarna och detektorspänningen för 7-ADD-parametern. Ställ in parametrarna så att 7-AAD-signalen för det ofärgade provet faller inom det första decenniet av logaritmningsskalan på punktdiagrammet. Börja förvärva röret med 7-AAD-färgade kärnor och definiera kärnpopulationerna genom att använda en gating-strategi baserad på (1) FSC-A/SCC-A och sedan FSC-H/SSC-H för storlek och granularitet, (2) FSC-H/FSC-A för dubblettdiskriminering och (3) SSC-A/7-AAD för 7-AAD-positiva kärnor (se figur 2A). Se till att strömmen och avböjningen är stabila. I sidoströmskameran, slå på testsorteringen, voltage PÅ, och bekräfta den korrekta droppsorteringen i ett 1.5 ml rör monterat på vänster sida. I fönstret Sorteringslayout väljer du den population som är av intresse, enligt definitionen i steg 2 (ovan). I Målhändelser väljer du tröskelvärdet i Kontinuerlig för att få minst 0,5 x 106 kärnor per prov. Under Precision väljer du 4-vägs renhet. När du är klar klickar du på Sortera och OK för att börja med kärnsorteringen. Kvalitetskontroll och räkning av renade kärnorOBS: Detta steg ska endast göras under pilotexperimentet för optimering av provberedningsstegen, med målet att testa renheten hos de sorterade kärnorna som ska laddas på 10X kromchipet. När protokollet är helt optimerat rekommenderas det inte att utföra detta kvalitetskontrollsteg i uppföljningsexperimenten för att undvika onödigt slöseri med insamlade kärnor som kan finnas tillgängliga i litet antal.Renhetskontroll med flödescytometriÖverför 10 μL av de sorterade kärnorna till ett nytt FACS-rör som innehåller 90 μL DPBS med 2 % värmeinaktiverat fetalt bovint serum (HI-FBS). Samla in och registrera eftersorteringsdata för att verifiera sorteringens renhet och genomförbarhet. Se till att minst 98 % av kärnorna visas i grinden av intresse, enligt definitionen i 4.2 (se figur 2B). Räkna de renade kärnornaCentrifugera sorterade kärnor i 5 minuter vid 500 x g och vid 4 °C och avlägsna försiktigt supernatanten helt med hjälp av en mikropipett. Återsuspendera i 100 μl utspädd kärnbuffert. Tillsätt 10 μL 0,4 % trypanblå till en tom 0,5 ml tub. Tillsätt 10 μl av de sorterade kärnorna och blanda 5 gånger genom pipettering. Räkna kärnor med hjälp av en automatiserad cellräknare enligt leverantörens rekommendationer. Justera kärnkoncentrationen till 3,5 x 106/ml, dvs 16 000 kärnor per 5 μL. Kvalitetskontroll av renade kärnor med mikroskopiOBS: Detta steg ska endast göras under pilotexperimentet för optimering av provberedningsstegen för att testa kvaliteten på kärnorna som ska laddas på 10X-kromchipet. När protokollet är helt optimerat rekommenderas det inte att utföra detta kvalitetskontrollsteg i uppföljningsexperimenten för att undvika onödigt slöseri med insamlade kärnor som kan finnas tillgängliga i litet antal.Se till att objektglas och täckglas är rena och dammfria. Tvätta och skölj vid behov täckglasen med absolut etanol och torka dem med luddfria våtservetter. Fördela 25 μL poly-l-lysin i de glasbrunnar som ska användas och inkubera i 10 minuter vid rumstemperatur (RT), skyddad från damm. Ta bort överskottet av poly-l-lysin och tillsätt 10 μl av den renade kärnsuspensionen. Inkubera i 5 minuter vid RT, skyddad från damm. Tillsätt en droppe monteringsmedium i varje brunn och undvik bubblor. Placera ett täckglas ovanpå de sådda brunnarna. Täck med pappersservetter och tryck fast täckglaset ordentligt för att ta bort överflödigt monteringsmedium. Var försiktig så att du inte flyttar täckglaset och rengör inte överflödigt monteringsmedium. Ta flera bilder med ett inverterat mikroskop med ljusfältsljus och en förstoring på minst 40x. Utför multiome-analys.Fortsätt omedelbart till Chromium Next GEM Single Cell Multiome ATAC + Gene Expression User Guide (CG000338 – Rev F)25. 2. Rening av kärnor från hematopoetiska stamceller och stamceller från musbenmärg (HSPC) OBS Detta protokoll beskriver rening av kärnor från tre undergrupper av benmärgens HSPC: härstamning-c-Kit+Sca-1+ hematopoetiska stamceller (HSC), härstamning-c-Kit+Sca-1-CD34+FcγR- vanliga myeloida stamceller (CMP) och härstamning- c-Kit+Sca-1-CD34+FcγR+ granulocyt-monocytprogenitorer (GMP). Använd latex- eller nitrilhandskar hela tiden under proceduren. Detta protokoll är en anpassning av 10X Genomics Shown Protocol – Nuclei Isolation for Single Cell Multiome ATAC + GEX-sekvensering (CG000365 – Rev C)27. Modifieringar har införts i det ursprungliga protokollet för att maximera kärnåtervinningen. Det rekommenderas starkt att ha två personer som utför experimentet, för att ha steg 1. till 3. (dvs. beredning av encellslösningen) utförs av en person, och steg 4 (dvs. beredning av sorteraren) utförs parallellt av en annan person. Eftersom protokollet är mycket tidskänsligt är det viktigt att minimera provbearbetningstiden genom att ha sorteraren redo så snart encellssuspensionen är förberedd. Beredning av reagenser och materialFyll en hink med is. Förbered FACS-bufferten: DPBS med 2 % HI-FBS-lösning (cirka 500 ml för 6 prover) och filtrera genom ett 0,2 μm filter. Förbered uppsamlingsmediet: DPBS med 10 % HI-FBS-lösning (500 μL per sample) och filtrera genom ett 0.2 μm filter. Isolering av benmärgscellerOffra möss med hjälp av protokoll som godkänts av institutionen. I detta experiment offrades mössen genom livmoderhalsluxation efter en överdos av ketamin/xylazin. Spraya mössens buk och bakben med 70% etanol. Använd en steril pincett och sax för att göra ett litet snitt i mitten av nedre delen av buken och öppna bukhinnan från basen av bakbenen till diafragman (tilläggsfigur 1). Gör ett extra snitt för varje bakben vinkelrätt mot den öppnade bukhinnan, ta sedan tag i vardera sidan av ett av dessa ytterligare snitt och dra isär det för att dra av huden från båda bakbenen förbi fotleden för att exponera musklerna i båda bakbenen (tilläggsfigur 1A). Rikta saxen längs ryggraden vid höftleden på ett bakben för att klippa ut benet utan att skära genom lårbenet (tilläggsfigur 1B, C). Upprepa samma sak för det andra benet. För att isolera lårbenet, skär ut det mesta av muskelvävnaden och håll sedan lårbenet och skenbenet i varje hand med fingertopparna vid leden (tilläggsfigur 1D, E). Vik försiktigt benet mot den naturliga böjningen för att flytta skenbenet från lårbenet (tilläggsfigur 1E) och klipp sedan försiktigt av bindväven med en sax för att separera lårbenet och skenbenet. Använd saxen med lätta vridande rörelser för att flytta ryggraden från lårbenets övre ände (tilläggsfigur 1E). Rengör det isolerade lårbenet med silkespapper för att ta bort kvarvarande muskler och bindväv. Förvara kallt på is i en 12-brunnars plattbrunn fylld med 2 ml DMEM (1x) + GlutaMAX-I. När alla lårben har samlats in, se till att muskel- och fibrösa vävnader är helt borttagna från benet. Skär inte upp benet för att (a) hålla märgen inuti steril och (b) undvika att förlora celler i brunnen. Använd följande steg för att spola cellerna från två lårben på en mus, anpassad från Haag och Murthy28. Förbered ett 1.5 ml och ett 0.5 ml rör. Tillsätt 150 μl FACS-buffert till 1,5 ml-röret, stick sedan ett hål i botten av 0,5 ml-röret med en 18 G-nål och sätt in 0,5 ml-röret i 1,5 ml-röret. Öppna den distala delen av varje lårben med en muskirurgisk sax (kompletterande figur 1F): Lås in den distala epifysen mellan bladen och tryck försiktigt samtidigt som du vänder saxen för att smidigt lossa den distala epifysen utan att skära upp benet hårt. Om det lyckas bör 4 utsprång vara synliga i den nu exponerade fysisänden (tilläggsfigur 1G). Montera de två lårbenen med den öppna änden nedåt i det förberedda 0,5 ml-röret som placeras inuti ett 1,5 ml-rör som innehåller FACS-buffert (tilläggsfigur 1H). Placera en 70 μm cellsil på ett 50 ml rör och förfukta silen med 2 ml FACS-buffert. För att spola benmärgen, centrifugera rören (öppna lock) vid 12 000 x g tills centrifugen når värdet 12 000 x g , stoppa sedan omedelbart centrifugen. Kontrollera att benmärgscellerna är pelleterade i 1,5 ml-röret och att lårbenen är vita (före cellspolning är de röda) (tilläggsfigur 1I). Kassera 0.5 ml-rören med de 2 lårbenen. Kassera supernatanten på 150 μl med en pipett. Återsuspendera pelleten med en mikropipett i 1 ml ammonium-klorid-kalium (ACK) lyseringsbuffert i 1-2 minuter vid RT för att lysera röda blodkroppar. Undvik längre inkubationstider eftersom de kan leda till minskad livskraft hos kärnförsedda celler. Överför till 50 ml-röret genom den förfuktade 70 μm cellsilen. Tillsätt 10 ml FACS-buffert för att späda ut ACK-lyseringsbufferten och stoppa därmed lyseringen. Centrifugera vid 400 x g i 5 minuter vid 4 °C. Återsuspendera i 10 ml FACS-buffert genom att först suspendera i 1 ml och sedan fylla på med 9 ml. Förbered cellerna för räkning enligt beskrivningen i 1.3.8. Räkna cellerna med hjälp av en automatiserad cellräknare enligt leverantörens rekommendationer. Den förväntas samla in cirka 40 miljoner celler från 2 lårben. Färgning av benmärg HSPCCentrifugera cellerna vid 400 x g i 5 minuter vid 4 °C och återsuspendera pelleten med en mikropipett i FACS-buffert till en slutlig koncentration av 1 x 107 celler/ml. Med en P1000-mikropipett överför du suspensionen till ett FACS-rör och filtrerar genom ett 35 μm cellsillock. Bered provrörsprover med en enda färgning för varje antikropp som anges i tabell 1 för att ställa in kompensationer av fluorokromer på cellsorteraren:Bered ett FACS-rör per antikropp och fyll rören med 200 μL PBS. Tillsätt 15 μL fluorokromkompensationskulor i varje FACS-rör med fluorkromkonjugerad antikropp. I FACS-rören för ofärgade och för levande/döda enfärgade celler, lägg till 500 000 celler i stället för pärlor. Tillsätt 1 μl av varje fluorokromkonjugerad antikropp (se tabell 1) i motsvarande FACS-rör. Tillsätt 0,5 μL levande/död fläck i levande/död FACS-rör med enkel fläck. Håll på isen i 15 minuter skyddad från ljus. Förbered blandning 1 och 2 enligt tabell 2.OBS: De antikroppsvolymer som anges i tabell 2 gäller för de antikroppar som refereras till i materialförteckningen. De måste optimeras för alla nya antikroppsreferenser eller ett annat parti av samma antikroppsreferens. Tillsätt 300 μL Mix 1 i provröret, sätt upp den och låt den stå i 15 minuter på is skyddad från ljus. Tillsätt 300 μL Mix 2 i provröret, sätt upp den och låt den stå i 20 minuter på is skyddad från ljus. Tillsätt 3 ml FACS-buffert till de enkelfärgade rören och de mixfärgade provrören. Centrifugera i 400 x g i 5 minuter vid 4 °C. Kassera försiktigt supernatanten med en mikropipett och återsuspendera pelleten i 500 μL FACS-bufferten. Förbered ett 1,5 ml rör förfyllt med 500 μl uppsamlingsmedium.OBS: Mix 1 är beredd i DPBS eftersom den innehåller Live/Dead-fläcken som påverkas avsevärt av HI-FBS. När cellerna är färgade av Live/Dead tillsätts Mix 2, som innehåller de fluorokromkonjugerade antikropparna resuspenderade i FACS-buffert innehållande HI-FBS. Det enda undantaget är anti-CD16/32-antikroppen som ingår i Antibody Mix 1 för att fungera som Fc-receptorblockerare som förhindrar icke-specifik bindning av de andra antikropparna som tillsätts i följande steg. Cellsortering med hjälp av en FACSOBS: Cellsortering kan utföras på en mängd olika cellsorterare, men här beskrivs proceduren för att använda instrumenten BD FACSAria Fusion eller BD FACSAria III. Det rekommenderas starkt att kalibreringen och installationen av cellsorteraren utförs under övervakning eller av en erfaren användare av instrumentet.Kalibrering av FACS-instrument: Se protokoll 1 steg 4.1. Uppsättning av FACS-instrument för cellsortering:Börja förvärva de ofärgade cellerna. Dessa används för att definiera framåt- och sidospridningen och detektorspänningen för varje fluorofor. Ställ in parametrarna så att den fluorescerande signalen för varje fluorofor faller inom det första decenniet av logaritmningsskalan på punktdiagrammet. Skaffa enfärgskontroller för att ställa in kompensationer manuellt (medianen för positiva och negativa populationer bör justeras) eller använd det automatiska beräkningsprogrammet (lutningsmätningar). Se till att kompensationskontrollerna matchar de experimentella fluorokromerna och detektorinställningarna. Spela in 10 000 händelser för celler och 5 000 händelser för pärlor. Använd provröret (dvs. flerfärgade celler) för att definiera cellpopulationer av intresse genom att använda grindstrategin som visas i figur 3A. Följ steg 4–6 (nedan). För att identifiera de tre benmärgs-HSPC:erna av intresse (HSC, CMP och GMP), starta grinden genom att använda storleken (FSC-A) och granulariteten (SSC-A) för att grinda på leukocyter, sedan FSC-H/FSC-A för att diskriminera dubbletter. Baserat på SSC-A/dödcellsmarkör, grinda levande celler. Använd Lineage/c-Kit för att välja celler som är ursprungsnegativa och uttrycker mellanliggande till höga nivåer av c-Kit. Genom c-Kit/Sca-1, grind på lineage-c-Kit+ Sca-1+ (LKS+) HSCs, en av de tre populationerna av intresse. Bland de myeloida stamfäderna (lineage-c-Kit+Sca-1-), använd FcγR/CD34 för att utesluta CD34-FcγR- megakaryocyt och erytroida progenitorer (MEP), samtidigt som CD34+ FcγR- CMP, samt CD34+FcγR+ GMP inkluderas i cellpopulationerna som ska sorteras. Se till att strömmen och avböjningen är stabila. I sidoströmskameran, slå på testsorteringen, voltage PÅ och bekräfta den korrekta droppsorteringen i ett 1.5 ml rör monterat på vänster sida. I fönstret Sorteringslayout väljer du den eller de populationer som är av intresse (t.ex. “LKS+” och “CD34+ myeloida förfäder” som visas i det här exemplet). Under Enhet väljer du 2 Tube. Under Precision väljer du Renhet. I Målhändelser väljer du Kontinuerlig för att sortera mellan 160 000 och 200 000 LKS+ och CD34+ myeloida förfäder. Tillsätt 500 μl FACS-buffert till cellsuspensionen och överför 1 ml av provet genom filtrering till ett nytt 35 μm-celltäckt FACS-rör med sil för att säkerställa att alla celler är i en enda suspension strax före upptaget. Detta eliminerar cellklumpar som kan täppa till instrumentet. När du är klar klickar du på Sortera och OK för att börja sortera. Justera flödeshastigheten för att hålla hastigheten under 10 000 händelser per sekund.OBS: Det förväntade förhållandet mellan LKS+ och CD34+ myeloida stamceller är 1:3 för en vuxen (8-12 veckor gammal) C57BL/6J honmus i steady state. De sorterade cellnumren nås vanligtvis inom 30 minuter efter sorteringen. Kvalitetskontroll och räkning av sorterade cellerOBS: Detta steg ska endast göras under pilotexperimentet för optimering av provberedningsstegen, med målet att testa renheten hos de sorterade cellerna som ska användas för kärnisolering. När protokollet är helt optimerat rekommenderas det inte att utföra detta kvalitetskontrollsteg i uppföljningsexperimenten för att undvika onödigt slöseri med utgångsmaterial som kan finnas tillgängligt i litet antal för isolering av kärnor.Renhetskontroll med flödescytometriÖverför 10 μL av de sorterade cellerna till ett nytt FACS-rör som innehåller 90 μL FACS-buffert. Samla in och registrera eftersorteringsdata för att verifiera sorteringens renhet och genomförbarhet. Se till att minst 95 % av cellerna visas i grinden av intresse, enligt definitionen i 3 – 6 och illustrerad i figur 3B. Isolering av kärnor från sorterade benmärgs-HSPCAnvänd protokollet “Low Cell Input Nuclei Isolation” i bilagan från 10X Genomics Shown Protocol – Nuclei Isolation for Single Cell Multiome ATAC + GEX-sekvensering (CG000365 – Rev C)27, med följande ändringar gjorda för att optimera kärnåtervinningen:Lystid: Kör ett pilotexperiment för det här protokollet för att identifiera den bästa lystiden för kärnisolering. Se till att uppnå en fullständig celllys samtidigt som du bibehåller intakta kärnor.OBS: Steg f i det ovan nämnda 10X Genomics-protokollet27 instruerar att “inkubera [i Lysis Buffer] i 3-5 minuter på is”. Under pilotexperimentet, testa minst 3 minuter, 4 minuter och 5 minuter och bedöm den återvunna kärnkvantiteten genom räkning och kvalitet genom flödescytometri och mikroskopiavbildning för att välja den optimala lystiden (se beskrivningen av dessa kvalitetskontrollkontroller nedan). För att spara reagens, byt ut den utspädda kärnbufferten mot PBS 0,04 % BSA i pilotexperimentet. För HSPC i benmärgen identifierades 3 min som den optimala lystiden. Cellcentrifugering: För alla cellsuspensionscentrifugeringar, centrifugera vid 300 x g i 7 minuter (i stället för 5 minuter i CG000365 – Rev C)27 vid 4 °C. Kärncentrifugeringar: Utför alla kärnsuspensionscentrifugeringar vid 500 x g i 5 minuter enligt CG000365 – Rev C27. Kärnuppsamling: I steg b, efter att ha suspenderat i 50 μl PBS 0,04 % BSA och överfört till ett 0,2 ml rör, tillsätt 50 μL PBS 0,04 % BSA till det ursprungliga röret och pipettblandningen för att samla upp eventuella överblivna celler. Överför till 0,2 ml röret för att nå en total volym på 100 μL. Hädanefter kommer den totala volymen att vara 100 μL i stället för 50 μL i protokollet. Justera stegen nedströms i enlighet med detta (t.ex. för steg d, ta bort 90 μL istället för 45 μL; för steg e, tillsätt 90 μL lysbuffert istället för 45 μL). För steg m återsuspenderas kärnpelleten i 12 μl utspädd kärnbuffert i stället för 7 μl. Räkna de isolerade kärnorna. Tillsätt 10 μL 0,4 % trypanblå och 8 μL PBS 0,04 % BSA i ett tomt 0,5 ml rör. Tillsätt 2 μl kärnor till röret och räkna kärnorna enligt beskrivningen i 1.3.8. Använd en automatiserad cellräknare enligt leverantörens rekommendationer. Renhetskontroll med flödescytometriOBS: Detta steg ska endast göras under pilotexperimentet för optimering av provberedningsstegen för att testa renheten hos kärnorna som ska laddas på 10X kromchipet. När protokollet är helt optimerat rekommenderas det inte att utföra detta kvalitetskontrollsteg i uppföljningsexperimenten för att undvika onödigt slöseri med insamlade kärnor som kan finnas tillgängliga i litet antal.Efter avslutad kärnisolering, överför 6 μL kärnresuspension till ett nytt FACS-rör förfyllt med 150 μL FACS-buffert. Tillsätt 3 μL 7-AAD och inkubera i 5 minuter på is. Samla in och registrera eftersorteringsdata för att verifiera sorteringens renhet och genomförbarhet. Se till att minst 95 % av kärnorna visas i grinden av intresse, enligt definitionen i protokoll 1 steg 4.2 (se figur 4). Kvalitetskontroll av renade kärnor med mikroskopi:OBS: Detta steg ska endast göras under pilotexperimentet för optimering av provberedningsstegen för att testa kvaliteten på kärnorna som ska laddas på 10X kromchipet. När protokollet är helt optimerat rekommenderas det inte att utföra detta kvalitetskontrollsteg i uppföljningsexperimenten för att undvika onödigt slöseri med insamlade kärnor som kan finnas tillgängliga i litet antal.Fortsätt enligt beskrivningen i steg 1.5.3. Utför multiome-analysFortsätt omedelbart till Chromium Next GEM Single Cell Multiome ATAC + Gene Expression User Guide (CG000338 – Rev F)25.