Cellemembranen, der tjener både som en fysisk barriere og en signalplatform, er afgørende for celleoverlevelse1. Gennem hele sin cellecyklus udsættes plasmamembranen (PM) for skader, såsom mekaniske 2,3,4,5 og kemiske6 stressinducerede skader. For at opretholde membranintegritet og sikre celleoverlevelse har cellen udviklet robuste plasmamembranreparationsmekanismer (PMR). Disse mekanismer afhænger af forskellige strategier, såsom cytoskelet reorganisering, membranfusion og membranudskiftningsstrategier 7,8,9,10,11, som alle er afhængige af rekruttering af specifikke proteiner. Især er medlemmer af annexinproteinfamilien blevet identificeret som nøgleproteiner forbundet med processerne i PMR 1,9,12,13,14,15,16. Efter PM-skade oplever cellen en tilstrømning af calciumioner (Ca2+), som udgør en umiddelbar trussel mod cellens overlevelse17. Som reaktion på Ca2+ tilstrømning binder annexinproteiner, som overvejende er placeret i cytosolen, til den indre folder af den beskadigede plasmamembran som en del af PMR-strategierne18. Annexin A2 (ANXA2) var et af de første medlemmer af annexin-familien, der blev forbundet med PMR i dysferlin-mangelfuld muskeldystrofi og blev foreslået at formidle reparation ved at fusionere intracellulære vesikler til PM nær skadestedet 5,19,20,21. Efterfølgende er flere funktioner blevet tilskrevet annexins22, og deres rolle i PMR har fået øget opmærksomhed i løbet af de sidste 20 år. Imidlertid er bilagenes nøjagtige rolle i PMR stadig ikke fuldt ud forstået 15,18,21,22.
Denne artikel foreslår en metode til at undersøge protein-membraninteraktion og membrandynamik på en kontrolleret og meget lokaliseret måde ved hjælp af en kombination af konfokal mikroskopi, optisk pincet og guldnanopartikler (AuNP’er). Denne metode muliggør kvantitativ undersøgelse af protein-, lipid- og små molekyleinteraktioner som reaktion på membranskader og Ca2+ tilstrømning. På trods af kompleksiteten og mangfoldigheden af komponenter, der er involveret i membranreparationsprocessen, er forenklede membransystemer, der efterligner plasmamembranen, blevet anvendt til at opnå en dybere mekanistisk forståelse af membrandynamik og annexinproteiners respons på membranforstyrrelser16. Gigantiske unilamellære lipidvesikler (GUV’er) blev valgt som modelmembransystem med en specificeret lipidsammensætning. GUV’erne blev genereret ved hjælp af gelassisteret hydreringsmetode, specifikt polyvinylalkoholgelhydrering, som beskrevet af Weinberger et al.23, hvilket muliggjorde effektiv indkapsling af annexiner i GUV’er.
Anvendelsen af nærinfrarød (NIR) laserbestråling på metalliske nanopartikler (NP’er) inducerer betydelig opvarmning af NP, hvilket gør det til en effektiv metode til at etablere en lokal varmekilde, der udnyttes i biomedicinske applikationer24. Metoden blev oprindeligt brugt til direkte at måle temperaturen omkring en enkelt AuNP i både 2D og 3D biomimetiske assays. I disse assays 25,26 blev de plasmoniske nanopartikler bestrålet på et understøttet lipiddobbeltlag eller optisk fanget nær GUV’er, der gennemgik en lokal termisk faseovergang ved lokal opvarmning, hvilket muliggjorde kvantificering og kontrol af den nøjagtige temperaturprofil omkring partiklen. Denne referencetemperaturprofil er blevet anvendt ved undersøgelse eller manipulation af biologiske prøver. Yderligere fremskridt i metoden har lettet induktionen af nanoskopiske porer i membraner27, hvilket muliggør vesikel og cellefusion 28,29. Andre undersøgelser har undersøgt opførsel af perifere membranproteiner i GUV’er29 og transmembranproteiner30 ved at skabe nye hybridvesikler, mens cellespecifik lægemiddellevering også er blevet undersøgt for at kontrollere og studere cellulære reaktioner eller genekspression 28,29,31,32,33. For nylig er metoden blevet brugt til at undersøge proteinresponser på membranskader 32,34,35.
Der findes flere metoder til at forstyrre plasmamembranen for at udforske cellulære reaktioner og membranreparation. Disse omfatter mikronålepunkteringer, mikroperlerystelser og celleskrabning, som alle kan forstyrre cellemembranen mekanisk 14,36,37. Kemisk induceret skade kan opnås ved at tilføje vaskemidler 5,38 eller bakterielle toksiner39,40, der destabiliserer lipiddobbeltlaget og genererer membranporer over plasmamembranen. Desuden er optisk inducerede skader ved kontinuerlige bølge- og pulserende lasere blevet anvendt til at studere PMR-komponenter, såsom annexinproteiner 5,14,21,41, i kombination med plasmoniske nanopartikler 42,43,44,45. På trods af effektiviteten af pulserende lasere med høj effekt kan de forårsage betydelige skader og skader på cellens indre langs strålebanen. Desuden mangler de detaljerede ændringer, der sker i det biologiske stof ved pulserende laserbestråling, og om det skaber en veldefineret pore, at blive undersøgt yderligere. En alternativ metode præsenteres i denne artikel, der anvender termoplasmonik til at inducere nanoskopiske huller i PM på en kontrolleret måde34,35 uden at forårsage væsentlig skade på de interne strukturer. Dette opnås ved at udsætte plasmoniske NP’er for en meget fokuseret NIR-laser, hvilket resulterer i en ekstremt lokaliseret temperaturstigning, der let kan nå temperaturer over 200 ° C, hvilket kan føre til små nanoskopiske eksplosioner 25,46,47. Denne proces kan styres ved at justere laserintensiteten samt størrelsen, formen og sammensætningen af NP’erne48. Ved at anvende denne teknik kan forskere undersøge proteiners rolle i PM-reparation i levende celler, hvilket kan hjælpe med at løse nogle af de ubesvarede spørgsmål vedrørende involvering af annexinproteiner i membranreparation uden at gå på kompromis med cellelevedygtigheden.
Den optiske fangst af plasmoniske nanopartikler er blevet veletableret af tidligere undersøgelser 25,49,50,51,52; yderligere indsigt i nanopartiklernes termoplasmoniske egenskaber 53,54,55 kan dog opnås i de supplerende materialer (supplerende fil 1). Den termoplasmoniske metode kan bruges til at skabe nanoskopiske huller i PM med det formål at studere de cellulære respons- og reparationsmekanismer. Mere præcist kan punkteringen opnås gennem optisk opvarmning af AuNP’er tæt på membranen, som vist i figur 1A og B. Denne lokaliserede punktering muliggør Ca2+ tilstrømning, som blev verificeret af en calciumsensor, hvilket aktiverede PMR-maskineriet. Til levende celleeksperimenter blev AuNP’er med en diameter på 200 nm immobiliseret på overfladen under cellen for at overvåge ANXA2’s rolle i PMR via konfokal mikroskopi. NIR-laseren (figur 1A, B) med en bølgelængde på 1064 nm bestråler AuNP og udnytter dens plasmoniske egenskaber (figur 1C), hvilket resulterer i betydelig lokal opvarmning (figur 1D) i det biologiske gennemsigtighedsvindue49, samtidig med at den forårsager minimal skade på selve cellen. Højtemperaturområdet omkring AuNP falder hurtigt med 30-40% i en afstand svarende til NP’s radius, som vist i figur 1E, hvilket giver mulighed for en ekstremt begrænset skade i alle tre dimensioner.
Supplerende dossier 1. Klik her for at downloade denne fil.
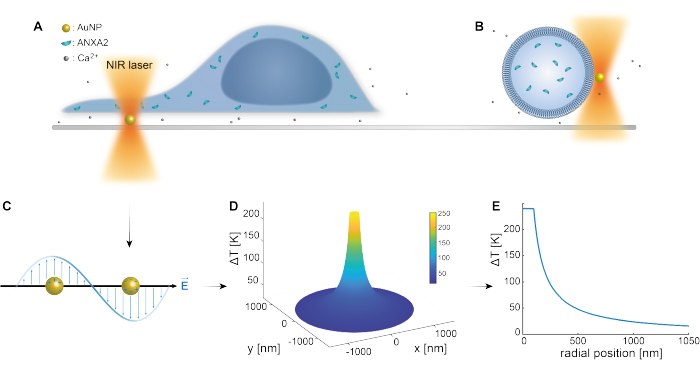
Figur 1: Skematisk oversigt over forsøgsmetoden. (A) ANXA-transfekterede celler er placeret oven på immobiliserede guldnanopartikler (AuNP’er) på overfladen, eller (B) gigantiske unilamellære vesikler (GUV’er) med indkapslet ANXA suspenderes i et medium indeholdende AuNP’er. C) En enkelt AuNP bestråles af den optiske NIR-fælde, hvor vekselvirkningen mellem det indkommende elektromagnetiske felt og ledningselektronerne fører til kollektiv svingning af elektroner i NP. (D) Denne proces resulterer i en meget begrænset, men signifikant temperaturstigning. For at estimere temperaturen på NP-overfladen anvendes Mie-teori, og en (E) temperaturprofil beregnes for en AuNP med en diameter på 200 nm og laserintensitet I = 6,36 x 108 W/cm2. Klik her for at se en større version af denne figur.
For at minimere den termiske effekt på cellemembranen bestråles AuNP’erne kun i ~ 1 sekund. Dette forårsager en forbigående og lokal udbrud af opvarmning, hvilket reducerer skaderne på proteiner, der typisk kræver mere tid at udfolde sig. Ved membranpunktering rekrutteres annexinproteiner i en brøkdel af et sekund, og inden for få sekunder dannes et annexinringlignende stillads omkring skadestedet (figur 2). Denne tilgang er også blevet anvendt til at undersøge involveringen af ANXA5 i både levende celler og modelmembraner16 i et forsøg på at kaste lys over hele reparationsprocessernes plan. Mens det primære fokus har været på den korrelerende rekruttering af forskellige annexinproteiner, er de biofysiske aspekter af reparationsmekanismen endnu ikke belyst.
For fuldt ud at gennemføre den foreslåede metode kræves tre nøglekomponenter: konfokal mikroskopi, optisk pincet og metal nanopartikler. Optisk pincet bruges til at fange AuNP’er, og deres konstruktion kan opnås ved at følge proceduren skitseret af Neuman et al.49. Men hvis opbygning af en optisk pincet viser sig at være for udfordrende, kan en tæt fokuseret NIR-laser bruges til at bestråle AuNP’er, der er immobiliseret under cellerne. Mens sfæriske AuNP’er blev valgt til denne protokol, kunne en række plasmoniske partikler med justerbare absorptionsspektre også anvendes til at opnå en meget lokaliseret temperaturgradient inden for NIR-regionen48.
Fluorescensbilleddannelse er nødvendig for at observere de fluorescerende mærkede proteiners rolle, og derfor kan total intern refleksionsmikroskopi (TIRF)56 betragtes som et alternativ til konfokal billeddannelse. Denne teknik tillader dog kun overfladebilleddannelse og ville ikke være kompatibel med modelmembranvesikeleksperimenterne. Derfor er både den optiske pincet og konfokalmikroskopet afgørende for den præcise lokalisering af nanopartiklen og detaljeret undersøgelse af lokalområdet omkring celleskaden. For effektivt at bestråle nanopartiklen med et diffraktionsbegrænset laserfokus er det nødvendigt at visualisere nanopartiklen. Dette kan opnås optimalt ved refleksionsmikroskopi, som er en standardbilleddannelsesfunktion i Leica konfokale mikroskoper. Men hvis refleksion eller spredningsbilleddannelse ikke er tilgængelig, kan alternative metoder, såsom den mindre effektive fluorescerende AuNP-mærkning, overvejes.
Sammenfattende har den meget kontrollerbare og lokaliserede termoplasmoniske metode, der præsenteres i denne undersøgelse, potentialet til at tjene som en fremragende platform til at undersøge de molekylære komponenter, der er involveret i cellulære reaktioner og PM-reparationsmekanismer i levende celler. Ud over at studere proteinresponset ved PM-skader kan denne tilgang også bruges til lokalt punktering af vesikler, hvilket muliggør en undersøgelse af proteinresponset i både protein-protein og protein-membrandynamik. Desuden giver denne metode mulighed for en kvantitativ analyse af interaktionerne mellem proteiner, lipider og små molekyler, når membraner forstyrres. Samlet set har disse fremskridt potentialet til at kaste lys over nogle af de uløste spørgsmål vedrørende de indviklede og komplekse plasmamembranreparationsmaskiner.
