A properly formed iBBB solidifies into a single translucent disc (Figure 3A). It is normal for the iBBB to detach from the surface onto which it was first pipetted after a few days. This cannot be avoided, but is not a major concern to the proper formation of the iBBB if care is taken when changing media to not accidentally aspirate the iBBB. After 24 h, evenly distributed, single cells can be identified under a brightfield microscope (Figure 3B). After 2 weeks, more distinct structures may be visible, although it is difficult to make out with definition (Figure 3C).
The quality of the iBBB is highly dependent on the quality of the differentiation of the input cells. We highly recommend plating some left-over cells in 2D monocultures to fix and stain for cell-type specific markers. iBBBs formed from endothelial cells that are over 70% positive for PECAM1 or VE-cadherin (Figure 2A), pericytes over 95% positive for PDGFRB (Figure 2B), and astrocytes over 95% positive for S100B and CD44 (Figure 2C) are best for successful iBBBs. The results from this quality check are the earliest indicator for high-quality iBBBs.
Physiologically relevant 3D structures should form after 2 weeks of self-assembly. Upon fixing and staining, we see evidence of tube-like structures that stain positive for the endothelial marker PECAM1, which is critical for tight junction formation (Figure 4A). The greatest variability in iBBB formation is the extent of microvasculature formation. In a "worst-case" scenario, the endothelial cell network appears more fragmented or does not extend throughout the iBBB, while in the "best-case" scenario, the vasculature is uniform and branches throughout the iBBB. Endothelial cell differentiation that is over 70% PECAM1-positive forms more consistent networks. Additionally, aquaporin-4, a protein expressed by astrocytes that localizes to the astrocyte endfeet, aligns with PECAM1 staining, indicating that astrocytes extend their endfeet to contact the endothelial cells (Figure 4B). Finally, we expect to see pericytes around the vasculature (Figure 4C).
The primary readout for cerebral amyloid angiopathy (CAA) in iBBBs is the presence of amyloid-β (Aβ). Aβ can be measured using targeted antibodies or by seeding fluorescently labeled Aβ to induce CAA phenotypes (Figure 5). Treatment of iBBBs with Aβ should increase Aβ staining intensity and area, as the cells of the BBB do not express a lot of endogenous Aβ protein. Alternatively, fixed samples can be stained with Thioflavin T to detect amyloid accumulation. Aβ levels are dependent on the genotype of the stem cells used to generate the iBBB, with some Alzheimer's disease and CAA-associated risk factors increasing the amount of Aβ accumulation and staining (Figure 5B, C)24.
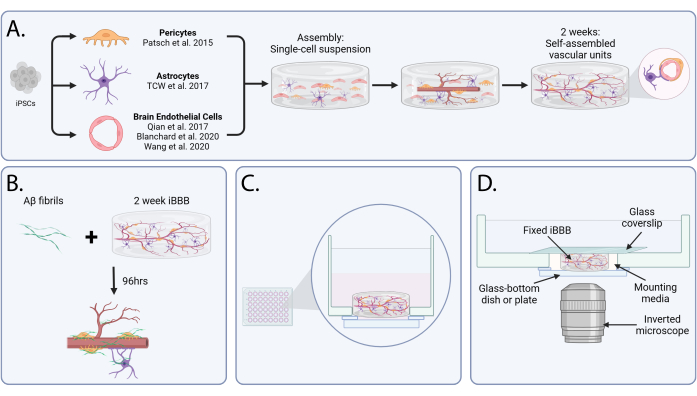
Figure 1: An in vitro blood-brain barrier to model cerebral amyloid angiopathy. (A) Schematic representation of iBBB assembly and maturation. After the differentiation of endothelial cells, astrocytes, and pericytes from induced pluripotent stem cells, cells are encapsulated in a gel matrix in a single-cell suspension. Over the course of 2 weeks, the cells self-assemble into vascular units, similar to the structures identified in vivo. (B) Schematic of the cerebral amyloid angiopathy assay. Amyloid-β is added to matured iBBBs for 96 h to induce Aβ aggregation. (C) Diagram showing the side view of an iBBB after seeding in a glass-bottomed well. (D) A graphic of an iBBB prepared for imaging on an inverted microscope. Abbreviations: BBB = blood-brain barrier; iPSCs = induced pluripotent stem cells; iBBB = an in vitro model of the human BBB derived from a patient iPSC-derived BBB model, which incorporates endothelial cells, pericytes, and astrocytes encapsulated in a 3D matrix; Aβ = Amyloid-β. Please click here to view a larger version of this figure.
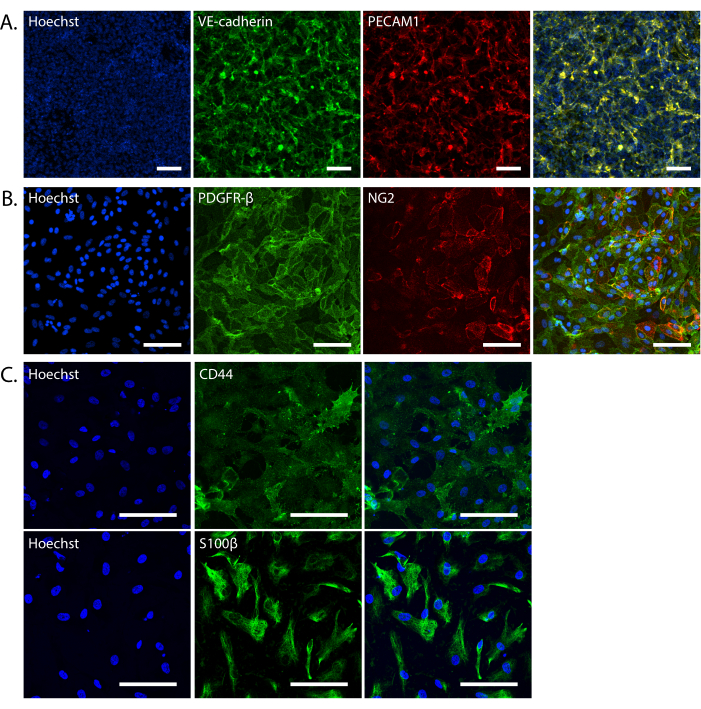
Figure 2: Validation of iPSC-derived iBBB cells. (A) Representative maximum intensity projection of endothelial cells in 2D monocultures stained with VE-cadherin (green) and PECAM1 (red). (B) Representative images of pericytes in 2D monocultures stained with PDGFRB (green) and NG2 (red). (C) Representative images of astrocytes in 2D monocultures stained with CD44 (top; green) and S100B (bottom; green). All nuclei are stained with Hoechst 33342. Images were taken on a Nikon Eclipse Ti2-E at 20x magnification (A,B) or a Leica Stellaris 8 at 40x magnification (C). All scale bars are 100 µm. Abbreviations: BBB = blood-brain barrier; iPSCs = induced pluripotent stem cells; iBBB = an in vitro model of the human BBB derived from a patient iPSC-derived BBB model, which incorporates endothelial cells, pericytes, and astrocytes encapsulated in a 3D matrix; VE-cadherin = vascular endothelial cadherin; PECAM1 = platelet and endothelial cell adhesion molecule 1; PDGFRB = platelet-derived growth factor receptor beta; NG2 = neuron glial antigen 2; S100B = S100 calcium-binding protein beta. Please click here to view a larger version of this figure.
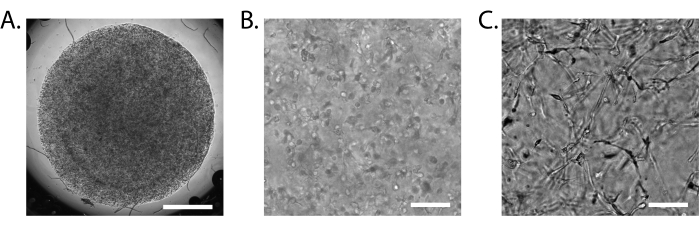
Figure 3: Assembly of the iBBB. (A) Brightfield image of a 15 µL iBBB 24 h after plating at 2x magnification. (B) Brightfield image of an iBBB 24 h after plating at 10x magnification. (C) Brightfield image of an iBBB 2 weeks after plating at 10x magnification. Scale bar = 1 mm (A), 100 µm (B,C). Images taken on an inverted Nikon Eclipse Ts2R-FL. Abbreviations: BBB = blood-brain barrier; iPSCs = induced pluripotent stem cells; iBBB = an in vitro model of the human BBB derived from a patient iPSC-derived BBB model, which incorporates endothelial cells, pericytes, and astrocytes encapsulated in a 3D matrix. Please click here to view a larger version of this figure.
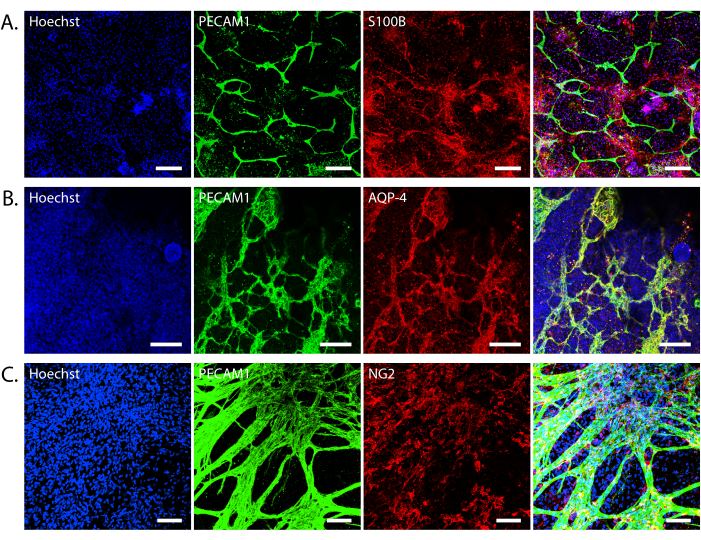
Figure 4: Cell interactions in the iBBB. Representative images of endothelial cells, pericytes, and astrocytes in the iBBB. (A) Endothelial cells (PECAM1) and astrocytes (S100β). (B) Endothelial cells (PECAM1) and astrocyte endfeet (AQP-4) co-localization. (C) endothelial cells (PECAM1) and pericytes (NG2). All nuclei are stained with Hoechst 33342. Confocal Z-stack images were taken on a Leica Stellaris 8 at 20x magnification (A,B) or a Nikon Eclipse Ti2-E at 20x magnification (C). Scale bars = 200 µm (A), 100 µm (B,C). Abbreviations: BBB = blood-brain barrier; iPSCs = induced pluripotent stem cells; iBBB = an in vitro model of the human BBB derived from a patient iPSC-derived BBB model, which incorporates endothelial cells, pericytes, and astrocytes encapsulated in a 3D matrix; PECAM1 = platelet and endothelial cell adhesion molecule 1; AQP-4 = aquaporin-4; NG2 = neuron glial antigen 2. Please click here to view a larger version of this figure.
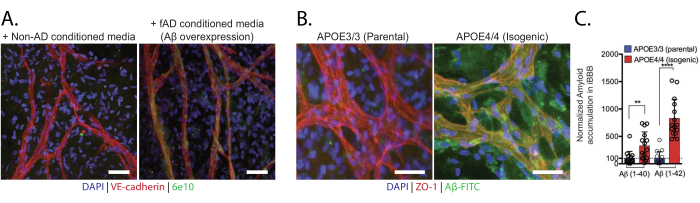
Figure 5: Cerebral amyloid angiopathy in vitro. (A) Non-AD iBBBs exposed to conditioned media from control or Aβ-overexpression neurons. 6e10 antibody recognizes Aβ. (B) Representative images of iBBBs from isogenic APOE3 and APOE4 cell lines treated with 20 nM Aβ-FITC1-42 for 96 h. (C) Quantification of amyloid in iBBBs from isogenic APOE3 and APOE4 cell lines treated with 20 nM Aβ-FITC1-40 or Aβ-FITC1-42.Scale bars = 50 µm (A), 10 µm (B). This figure was adapted from Blanchard et al24. Abbreviations: BBB = blood-brain barrier; iPSCs = induced pluripotent stem cells; iBBB = an in vitro model of the human BBB derived from a patient iPSC-derived BBB model, which incorporates endothelial cells, pericytes, and astrocytes encapsulated in a 3D matrix; Aβ = Amyloid-β. APOE = ApolipoproteinE. Please click here to view a larger version of this figure.
| Markers | Company | Catalog Number | Dilution | |
| Endothelial Cells | PECAM1 (CD31) | R&D Systems | AF806 | 1:500 |
| VE-cadherin (CD144) | R&D systems | AF938 | 1:500 | |
| ZO-1 | Invitrogen | MA3-39100-A488 | 1:500 | |
| Pericytes | PDGFRβ | R&D Systems | AF385 | 1:500 |
| NG2 | Abcam | ab255811 | 1:500 | |
| Astrocytes | S100β | Sigma-Aldrich | S2532-100uL | 1:500 |
| CD44 | Cell Signaling Technology | 3570S | 1:400 | |
| AQP-4 | Invitrogen | PA5-53234 | 1:300 | |
| GFAP | ||||
| ALDH1L1 | ||||
| EAAT1 | ||||
| EAAT2 | ||||
| Amyloid-β | 6e10 | Biolegend | SIG-39320 | 1:1,000 |
| Thioflavin T | Chem Impex | 22870 | 25 µM |
Table 1: Recommended cell markers for differentiation quality checks. Cell markers for the different cell types of the BBB that can be used to check the quality of the differentiations and to identify the cells in the formed iBBB. The markers used in this paper are bolded.
