Quantification of Autoreactive Antibodies in Mice upon Experimental Autoimmune Encephalomyelitis
Summary
Experimental autoimmune encephalomyelitis (EAE) is an animal model of multiple sclerosis (MS), which shares with the human disease a robust humoral autoimmune response. Here, we report a simple and flexible ELISA protocol to quantify autoantibodies in the serum of EAE immunized mice.
Abstract
Experimental autoimmune encephalomyelitis (EAE) is a disease model that recapitulates the autoimmune disorder multiple sclerosis (MS) at histopathological and molecular levels. EAE is induced by immunizing experimental animals via subcutaneous injection of short myelin peptides together with specific adjuvants to boost the immune response. Like the human counterpart, EAE mice develop demyelinating lesions, immune cell infiltration into the central nervous system (CNS), glia activation and neuronal injury. A consistent body of evidence also supports a mechanistic role for B cell dysfunction in the etiology of both MS and EAE. B cells can serve as antigen-presenting cells as well as a primary source of pro-inflammatory cytokines and autoantibodies. In EAE, antibodies are generated against the myelin peptides that were employed to induce the disease. Such autoantibodies have been shown to mediate either myelin loss or pathogenic T cell reactivation into the CNS. This article describes an efficient ELISA-based protocol to quantify autoantibodies in the serum of C57BL/6J mice immunized with the myelin oligodendrocyte glycoprotein 35-55 (MOG35-55) peptide. The proposed method serves as a powerful tool to investigate the specificity and magnitude of the aberrant humoral response in the context of autoimmune demyelination.
Introduction
Multiple sclerosis (MS) is a chronic autoimmune disease of the central nervous system (CNS) characterized by focal infiltration of immune cells into the brain parenchyma, breakdown of myelin sheaths wrapping axons, glia activation, and neuronal loss1. In addition to the well-established role of pathogenic T cells, multiple lines of evidence have highlighted the involvement of B cells in mediating the autoimmune response against the CNS. B cells undergo clonal expansion in the MS brain and antibodies against myelin components have been detected within demyelinated lesions2,3. The selective activation of peripheral B cells at disease onset has been recently documented, suggesting a putative role for this immune cell compartment in disease initiation as well4. The success of B cell-depleting therapies such as anti-CD20 monoclonal antibodies further corroborates the mechanistic connection between aberrant B cell functioning and autoimmune demyelination5,6. From a molecular standpoint, B cells can contribute to disease via autoantigen presentation, pro-inflammatory cytokine secretion, and autoreactive antibody production.
Multiple animal models have been developed to recapitulate specific features of the complex MS phenotype. Among them, experimental autoimmune encephalomyelitis (EAE) is the most widely used in vivo paradigm and relies on the immunization of experimental animals with short peptides derived from myelin proteins such as myelin oligodendrocyte glycoprotein (MOG) and myelin basic protein (MBP)7. EAE immunized animals develop a demyelinating pathology that resembles MS in many aspects, including a robust humoral response against the encephalitogenic peptide8. For this reason, EAE studies have been instrumental in dissecting the function of B cells and autoantibodies in the context of disease. For instance, it was demonstrated that MOG-specific antibodies isolated from MS patients can aggravate the clinical course in EAE models9. Notably, the proline residue at position 42 in human MOG was shown to be critical for determining autoantibody pathogenicity10. More recently, MOG-specific autoantibodies have been found to promote disease not only by mediating myelin loss but also via boosting the reactivation of autoreactive T cells within the CNS11.
Considering the importance of antibody responses in CNS autoimmunity, this article presents an ELISA-based protocol to efficiently measure the serum levels of autoreactive antibodies in C57BL/6J mice EAE immunized with MOG35-55 peptide. In the first part of the protocol, the method to collect serum via intracardiac puncture will be described. Subsequently, the procedures to set up the ELISA assay and acquire the data will be detailed. Lastly, data analysis and interpretation will be discussed.
Protocol
All procedures involving mice were performed in compliance with experimental guidelines approved by the East Carolina University Institutional Animal Care and Use Committee (IACUC). Wildtype C57BL/6J female mice between 8-10 weeks of age were used in this study. The animals were obtained from a commercial source (see Table of Materials). EAE was induced following previously published reports12,13,14.
1. Serum collection
- Euthanize the experimental mouse by CO2 asphyxiation or isoflurane overdose at the required time point (following institutionally approved protocols) after inducing EAE via MOG35-55 immunization.
NOTE: The total number of mice and time points for serum collection can vary according to the specific experiment. - After the absence of vital signs is confirmed by toe or tail pinch, place the mouse in a dorsal recumbency position on a dissection tray and affix the limbs in position with pins or tape. Orient the mouse body to the LED light source (see Table of Materials) to illuminate the thorax of the animal.
- Spray the mouse fur with 70% ethanol and make a midline incision of 3-4 cm along the abdomen from the pelvis to the xiphoid using the dissector scissors, taking care to avoid any organs and major vessels (Figure 1A-C). To facilitate the procedure, use the forceps to grab the skin over the xiphoid process.
- Cut through the diaphragm laterally and then cut the rib cage anteriorly on both lateral edges using the spring scissors, stopping before reaching the sternum at the midline (Figure 1D). Fold the rib flap that was created over the mouse head to expose the heart (Figure 1E).
NOTE: Cutting the sternum should be avoided as this will damage the main thoracic arteries laying adjacent to the bone and reduce the volume of blood that can be collected. - Insert a 25 G needle connected to a 1 mL syringe into the left ventricle and collect the blood by gently pulling back the plunger (Figure 1F). To facilitate the needle puncture, gently brace the heart against a pair of forceps.
NOTE: If the blood does not appear immediately into the syringe, the needle should be slowly rotated, and a different angle should be tested. However, efforts should be made to place the needle properly on the first attempt to avoid creating unnecessary holes in the heart where blood can leak out. - Transfer the blood into a sterile 1.5 mL tube and allow it to clot for 30 min at room temperature. Remove the clot by centrifugation at 2000 × g for 20 min at 4 °C. Collect the supernatant, which represents the serum fraction, and store single-use aliquots into cryogenic tubes at -80 °C for future testing.
2. ELISA assay
- Resuspend the lyophilized MOG35-55 peptide (see Table of Materials) in water to obtain a 10 mg/mL stock. Before starting the experiment, dilute the stock to 10 μg/mL in coating buffer (see Table of Materials) and pipette 100 μL/well of the final solution into a 96-well plate. Seal the plate with an adhesive film to avoid evaporation and incubate the plate overnight at 4 °C.
NOTE: At least 2 wells should be calculated for each sample and 2 additional wells should be also included for a blank control. - In parallel, coat the same number of wells with 100 μL/well of bovine serum albumin (BSA) dissolved in coating buffer at 10 μg/mL concentration. These additional wells will serve as background controls.
NOTE: It is recommended to coat with BSA the controls wells since the coating and blocking buffers have different compositions. Please note that other encephalitogenic peptides (such as PLP139-151 or MBP84-104) could be used as irrelevant antigens in alternative to BSA. - The day after, wash the plate 3 times with 200 μL/well of phosphate buffer saline supplemented with 0.05% Tween 20 (PBS-T). Subsequently, add 100 μL/well of a blocking solution made of 3% BSA in PBS (without Tween 20) and incubate the plate sealed for 1 h at 37 °C in a hybridization oven.
- Wash the plate 3 times with 200 μL/well of PBS-T. Dilute each serum sample 1:100 in blocking solution and add 100 μL/well to both MOG35-55 and BSA coated wells. Add the same volume of blocking solution to the wells designated as blanks. Incubate the plate sealed for 2 h at room temperature, with constant shaking (250 rpm).
- Wash the plate 3 times with 200 μL/well of PBS-T. Dilute the HRP-conjugated secondary antibody (1:2000, see Table of Materials) in a solution made of 0.2% BSA in PBS-T and add 100 μL/well to all wells. Incubate the plate sealed for 1 h at room temperature, with constant shaking (250 rpm).
- Wash the plate 3 times with 200 μL/well of PBS-T. Add 100 µL/well of 3,3',5,5' tetramethylbenzidine (TMB) substrate (see Table of Materials) to all the wells and incubate in the dark for 1-5 min, monitoring the development of a blue color.
- Stop the reaction by adding 100 µL/well of stop solution (see Table of Materials) and measure the optical density (OD) in each well using a plate reader set at a wavelength of 450 nm.
NOTE: The TMB substrate should be equilibrated at room temperature for 30-60 minutes before adding it to the wells.
- Stop the reaction by adding 100 µL/well of stop solution (see Table of Materials) and measure the optical density (OD) in each well using a plate reader set at a wavelength of 450 nm.
3. Data analysis
- Populate an Excel spreadsheet with the OD values read from the plate. Average the OD values obtained from duplicate wells (both MOG35-55 and BSA coated) for each sample.
- For each sample, subtract the mean value of the BSA-coated wells from that of MOG35-55 coated wells. The resulting background corrected values will be proportional to the concentration of anti-MOG35-55 antibodies in the different serum samples.
NOTE: Blank wells should result in corrected values around 0. - Apply a non-parametric statistical test such as the Mann-Whitney U test to compare the average OD values between two experimental conditions. Include at least 3 independent samples for each condition.
Representative Results
To demonstrate the robustness of the present ELISA assay, the method was tested on serum samples isolated from a cohort of C57BL/6J female mice at 20 days post-immunization (dpi) with 100 μg of MOG35-55 peptide in complete Freund's adjuvant (CFA) following a validated EAE induction protocol12,13,14. The animals also received 400 ng of pertussis toxin on day 0 and 2. Serum samples from mock immunized animals with everything but without the peptide served as negative controls. All EAE mice developed the first signs of disease between 11-14 dpi and reached by 20 dpi an average score of 2.6 ± 0.6 (standard error, SE) on a 0-6 scale (0, no signs; 1, decreased tail tone; 2, mild monoparesis or paraparesis; 3, severe paraparesis; 4, paraplegia; 5, quadriparesis; and 6, moribund or death)12. As expected, the EAE samples displayed significantly higher OD values compared to control samples (Figure 2). These data confirm the presence of a robust IgG immunoglobulin response against the MOG peptide at the peak of disease.
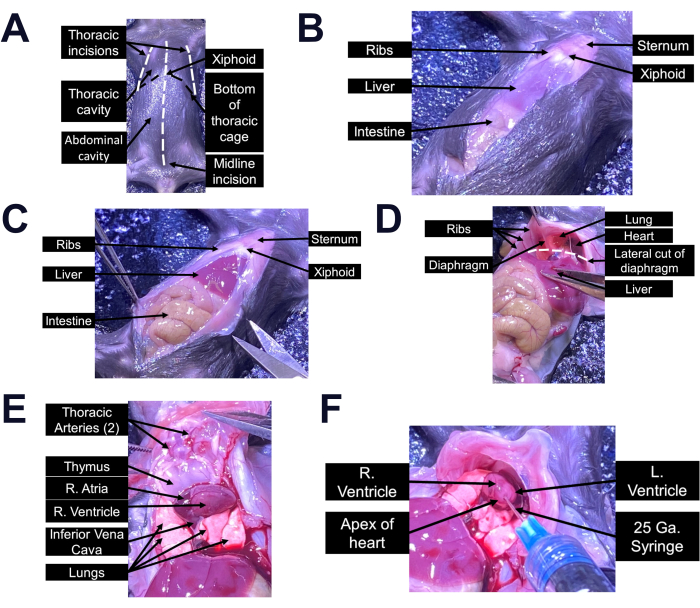
Figure 1: Cardiac puncture procedure. (A–E) Representative images of the thoracotomy procedure to access the heart in adult mice. (F) Representative image of the correct procedure for needle insertion into the left ventricle of the mouse heart. Relevant anatomical structures are indicated in each panel to facilitate the dissection. Please click here to view a larger version of this figure.
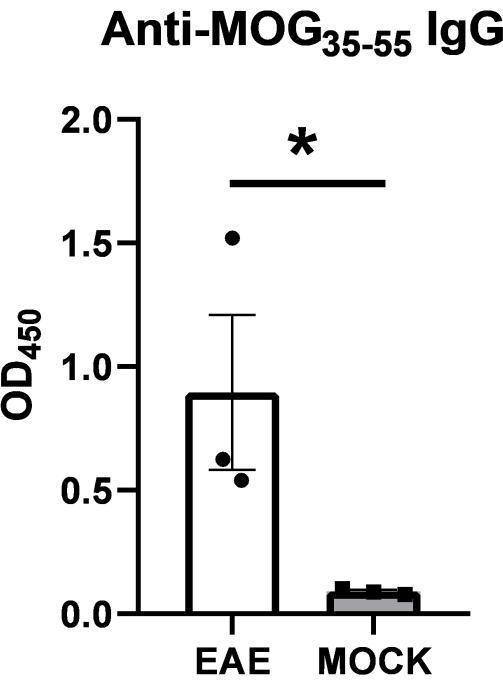
Figure 2: MOG peptide autoantibody ELISA. Serum levels of anti-MOG35-55 IgG immunoglobulins were tested in EAE and mock immunized mice at 20 days post-immunization (dpi) using the reported ELISA protocol. Consistently higher OD values were detected in EAE samples compared to controls. Data are presented as means ± SE (N = 3 per group). Differences between groups were assessed by one-tailed Mann-Whitney U test, *P ≤ 0.05. Please click here to view a larger version of this figure.
Discussion
Here, a simple and efficient ELISA-based protocol was reported to accurately quantify the humoral response in a relevant animal model of MS pathology. This method has been recently employed to describe the novel role of the ataxin-1 protein in controlling the serum levels of autoantibodies in the MOG35-55/C57BL6J EAE paradigm12. In this regard, a number of factors should be taken into consideration, in order to obtain consistent and biologically meaningful results with this method.
First, it is crucial to collect high quality serum samples and avoid hemolysis. Hemoglobin and other intracellular components released from red blood cells in heavily hemolyzed specimens can interfere with the antigen-antibody reaction and relative color measurement in the ELISA assay. To prevent hemolysis, it is preferable to avoid high negative pressure in the syringe during blood collection as this can cause erythrocytes to rupture.
Second, it is important to dilute the serum samples to obtain OD values within the linear portion of the absorbance curve. Our recommended dilution factor (1:100) was calibrated on the entity of the EAE pathology induced in our mouse strains. Considering the intrinsic level of heterogeneity of this in vivo model, it is highly recommended to perform preliminary titration experiments to identify the ideal dilution factor for the specific EAE pathology that is tested. Likewise, the incubation times with the TMB substrate may be adjusted to avoid the saturation of the chromogenic signal and maintain the OD values within the 0-2 range.
Third, the corrected OD values obtained in this protocol are compatible with a semi-quantitative analysis of autoantibody serum titers between different experimental conditions or for kinetic studies along disease progression. If an absolute quantification of the autoantibody levels is needed, a calibration curve must be implemented in the ELISA design. Multiple antibodies against MOG peptide are commercially available and can be used as reliable standards to calculate the concentration of MOG-specific antibodies in the serum of EAE mice. To capture effects of different magnitude, it is recommended to include a wide range of dilutions for the calibrating antibody (ranging from pg/mL to μg/mL concentrations).
Lastly, this protocol is extremely flexible and can be easily adapted to accommodate different research needs. For instance, the coated encephalitogenic peptide can be varied to assess the autoantibody response in other EAE paradigms. Furthermore, different secondary antibodies can be employed to measure the levels of distinct immunoglobulin classes. To this end, this protocol successfully demonstrated that ataxin-1 controls both IgG and IgM classes of MOG peptide-specific antibodies12. Lastly, it could be envisioned a possible use for detecting autoantibodies in the blood of MS patients. However, substantial differences exist between EAE and MS diseases. While the former is artificially induced via boosting the immune response against a well-defined autoantigen with strong adjuvants, the latter arises spontaneously and no unambiguous autoantigens have been identified yet. Recent evidence suggests the possible involvement of molecular mimicry with viral proteins15. Hence, caveats should always be considered in directly translating observations collected in this animal model to the human disease.
In summary, the present protocol represents an easy and convenient alternative to commercial ELISA kits and could be efficiently combined with other methods such as cell-based assays (CBAs) for the precise estimation of autoantibody release in the context of autoimmune demyelination.
Declarações
The authors have nothing to disclose.
Acknowledgements
This study was supported by the National Institutes of Health (R03NS131908) and the Department of Defense through the Multiple Sclerosis Research Program under Award No. W81XWH-22-1-0517. Opinions, interpretations, conclusions and recommendations are those of the author and are not necessarily endorsed by the Department of Defense. This study was also supported by East Carolina University startup funds.
Materials
| 1 mL syringes | BD Biosciences | 309628 | |
| 1.5 mL microcentrifuge tubes | Fisher | 05-408-129 | |
| 25 G needles | BD Biosciences | 305122 | |
| 3,3',5,5'-tetramethylbenzidine (TMB) substrate | Thermo Fisher | N301 | Store at 4 °C |
| Adhesive seals | Thermo Fisher | AB0558 | |
| Bovine serum albumin (BSA) | Sigma | A7906 | Store at 4 °C |
| C57BL/6J female mice | The Jackson Laboratory | 000664 | Animals between 8-10 weeks of age should be used for EAE experiments |
| Cryogenic tubes | Fisher | 10-500-25 | |
| Dissection tray | Fisher | S111022 | |
| Dissector scissors | Fine Science Tools | 14082-09 | |
| ELISA coating buffer | BioLegend | 421701 | Store at 4°C |
| Excel software | Microsoft | Analysis spreadsheet | |
| Forceps | Fine Science Tools | 11152-10 | |
| Goat Anti-Mouse IgG, Human ads-HRP | SouthernBiotech | 1030-05 | Store at 4 °C |
| LED light source | Fisher | AMPSILED21 | |
| Microplate reader | Fisher | 14-377-575 | |
| Molecular biology grade water | Corning | 46-000-Cl | |
| Mouse MOG35-55 peptide | EZBiolab | cp7203 | Store at -80 °C |
| Multichannel pipette | Axygen | AP-12-200-P | |
| Noyes spring scissors | Fine Science Tools | 15011-12 | |
| Nunc MaxiSorp 96-well plates | BioLegend | 423501 | |
| Orbital shaker | Fisher | 88-861-023 | |
| Oven | VWR | 445-0024 | |
| Phosphate buffer saline (PBS) | Thermo Fisher | 14190144 | |
| Refrigerated tabletop centrifuge | Thermo Fisher | 75002441 | |
| Stop solution | Thermo Fisher | N600 | |
| Tween 20 | Bio-Rad | 1706531 |
Referências
- Reich, D. S., Lucchinetti, C. F., Calabresi, P. A. Multiple sclerosis. N Engl J Med. 378 (2), 169-180 (2018).
- Baranzini, S. E., et al. B cell repertoire diversity and clonal expansion in multiple sclerosis brain lesions. J Immunol. 163 (9), 5133-5144 (1999).
- Genain, C. P., Cannella, B., Hauser, S. L., Raine, C. S. Identification of autoantibodies associated with myelin damage in multiple sclerosis. Nat Med. 5 (2), 170-175 (1999).
- Ma, Q., et al. Specific hypomethylation programs underpin b cell activation in early multiple sclerosis. Proc Natl Acad Sci U S A. 118 (51), e2111920118 (2021).
- Hauser, S. L., et al. Ocrelizumab versus interferon beta-1a in relapsing multiple sclerosis. N Engl J Med. 376 (3), 221-234 (2017).
- Montalban, X., et al. Ocrelizumab versus placebo in primary progressive multiple sclerosis. N Engl J Med. 376 (3), 209-220 (2017).
- Didonna, A. Preclinical models of multiple sclerosis: Advantages and limitations towards better therapies. Curr Med Chem. 23 (14), 1442-1459 (2016).
- Steinman, L., Zamvil, S. S. How to successfully apply animal studies in experimental allergic encephalomyelitis to research on multiple sclerosis. Ann Neurol. 60 (1), 12-21 (2006).
- Khare, P., et al. Myelin oligodendrocyte glycoprotein-specific antibodies from multiple sclerosis patients exacerbate disease in a humanized mouse model. J Autoimmun. 86, 104-115 (2018).
- Marta, C. B., Oliver, A. R., Sweet, R. A., Pfeiffer, S. E., Ruddle, N. H. Pathogenic myelin oligodendrocyte glycoprotein antibodies recognize glycosylated epitopes and perturb oligodendrocyte physiology. Proc Natl Acad Sci U S A. 102 (39), 13992-13997 (2005).
- Flach, A. C., et al. Autoantibody-boosted t-cell reactivation in the target organ triggers manifestation of autoimmune cns disease. Proc Natl Acad Sci U S A. 113 (12), 3323-3328 (2016).
- Didonna, A., et al. Ataxin-1 regulates b cell function and the severity of autoimmune experimental encephalomyelitis. Proc Natl Acad Sci U S A. 117 (38), 23742-23750 (2020).
- Didonna, A., et al. Sex-specific tau methylation patterns and synaptic transcriptional alterations are associated with neural vulnerability during chronic neuroinflammation. J Autoimmun. 101, 56-69 (2019).
- Ma, Q., Matsunaga, A., Ho, B., Oksenberg, J. R., Didonna, A. Oligodendrocyte-specific argonaute profiling identifies micrornas associated with experimental autoimmune encephalomyelitis. J Neuroinflammation. 17 (1), 297 (2020).
- Lanz, T. V., et al. Clonally expanded b cells in multiple sclerosis bind ebv ebna1 and glialcam. Nature. 603 (7900), 321-327 (2022).
