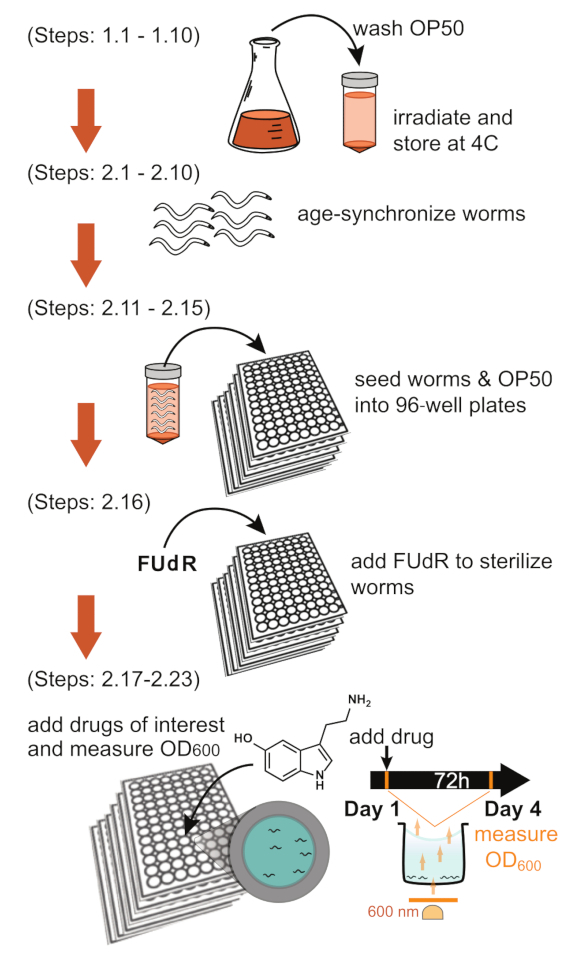
Figure 1: Schematic of the bacterial clearance assay. Graphical outline of the main sections of the protocol (Top-to-bottom): Preparing bacteria, synchronizing the worm population, seeding in 96-well plates, sterilizing worms, adding drugs, measuring OD600 on day 1 and day 4. A detailed description of these specific steps is indicated to the left of each depiction. Abbreviations: OD = optical density; FUdR = fluorodeoxyuridine. Please click here to view a larger version of this figure.
1. Bacteria preparation
NOTE: This section details the preparation of the feeding bacteria used in this assay. The specific E. coli strain used in the assay is called OP50. OP50 is the most widely used strain to feed C. elegans. The OP50 strain used is Carbenicillin/Ampicillin-resistant, which prevents cross-contamination of the worm culture with other bacteria9.
- Prepare the OP50 4-5 days in advance, and x-ray irradiate a day prior to usage. Ensure that all materials encountering OP50 are sterile29,30.
- Day -8: Wednesday (week 1):
- Prepare the preinoculum by inoculating 5 mL of LB with 100 µg/mL Ampicillin and 0.1 µg/mL Amphotericin B, and a single OP50 colony and incubate for approximately 6 h at 37 °C in a bacterial shaker. Inoculate early to allow sufficient growth for the culture to become cloudy.
- Once the culture is cloudy, dilute the preinoculum culture of OP50 by 1:2,000 in 250 mL of TB containing 100 µg/mL Ampicillin. Incubate the culture overnight in a bacterial shaker for 17-19 h at 37 °C.
NOTE: The culture should not be allowed to grow longer than 19 h.
- Day -7: Thursday (week 1)
- After the 17-19 h incubation, transfer the OP50 liquid culture into a sterile centrifugation tube and centrifuge for 15 min at 3,100 × g in a tabletop centrifuge at 4 °C.
- Discard the supernatant, resuspend the OP50 pellet with sterile water, and re-centrifuge. Repeat this wash 2x.
- After the second wash, discard the supernatant, resuspend the OP50 pellet in sterile water to a volume of 50 mL and transfer it to a preweighed 50 mL conical tube. Pellet the OP50 by centrifugation for 20 min at 3,100 × g in a tabletop centrifuge at 4 °C.
- After the third wash, carefully remove all the remaining supernatant, ensuring no water remains in the tube. Calculate the weight of the pellet by subtracting the weight of the empty centrifugation tube from the weight of the centrifugation tube with the pellet.
NOTE: Pellet weights typically range from 3-3.5 g. - Thoroughly resuspend the OP50 pellet in the S-complete buffer to a concentration of 100 mg/ mL, ensuring no clumps are present.
- Ensure that the concentration of OP50 at 100 mg/mL corresponds to 2 × 1010 bacteria/mL. If the relationship between the optical density and the number of bacteria per mL is known, determine the bacterial concentration per mL spectrophotometrically. If necessary, use S-complete buffer to adjust the concentration of the OP50 feeding solution to 2 × 1010 bacteria/mL.
- Test the OP50 on an agar plate to determine if it has been contaminated.
- Store the OP50 suspended in S-complete buffer at 4 °C.
- Day -3: Monday (week 2)
- Kill the bacteria via X-ray irradiation to prevent bacterial growth during the assay.
2. Synchronous worm culture preparation
NOTE: This section discusses the preparation of a synchronous worm population. All materials that come into contact with the worm populations are sterile. All plates are kept at 20 °C unless otherwise noted.
- Day -6: Friday (week 1), 4:00 p.m.
- Transfer the animals to a fresh plate.
- Take an NGM plate on which most of the worm population consists of starved L1 larvae.
NOTE: A mixed population will also produce results. Using L1 worms that have been starved for a long time may affect the feeding behavior in the current or future generations, as starvation can leave epigenetic marks for at least three generations. - Wash the worms off with no more than 1 mL of sterile water. Aliquot ~300 µL of the washed-off worm population onto NGM plates seeded with concentrated OP50 (~300 mg/mL). Allow the plates to dry under either a plate drier or a Bunsen burner. Incubate the plates at 20°C until much of the population contains gravid adults (Monday).
NOTE: This time frame is optimized for a wild-type N2 worm population and may vary from strain to strain. Worms with development delays should be taken into account and be chunked earlier and/or seeded earlier so that all strains tested will reach adulthood simultaneously.
- Take an NGM plate on which most of the worm population consists of starved L1 larvae.
- Transfer the animals to a fresh plate.
- Day -3: Monday (week 2) 10:00 a.m.
- Establishing a synchronous population
CAUTION! Bleach is corrosive to skin and eyes. Handling it requires gloves and safety glasses.- Collect worms by washing them off the plate with up to 10 mL of sterile water. Transfer this worm/water solution into a 15 mL conical tube.
- Wash the worms by gravity sink on the bench top for approximately 4 min (conversely, wash by centrifugation for 2 min in a tabletop centrifuge at 310 × g.
- Discard the supernatant through aspiration using a small tip and add up to 15 mL of sterile water. Repeat 3x.
- Remove the supernatant and add 5 mL of a freshly prepared bleach/NaOH solution (1.8 mL of household bleach, 0.5 mL of 10 N NaOH, 7.7 mL of dH2O).
- Incubate for 5 min at room temperature or until the worms break open. Vortex gently every minute for at least 10 s. Monitor the progress under the dissecting microscope.
NOTE: The time necessary to break open the worms may vary from strain to strain. Leaving the worms in the bleach solution for too long may result in nonviable eggs. - Add M9 buffer to neutralize the reaction once all the adults break open or dissolve (final volume to be 15 mL) and centrifuge for 2 min at 1,300 × g.
- Wash the eggs 3x with 13 mL of M9 buffer.
- Wash the eggs once with 10 mL of S-complete buffer by centrifuging for 2 min at 1,300 × g.
- Aspirate the supernatant, add 10 mL of S-complete, and transfer the suspension to a fresh 15 mL conical tube. Gently rotate the tube at room temperature overnight on a nutator or similar device.
- Optional: If adult carcasses are present after final centrifugation in S-complete buffer, filter the worm solution through a 40 µm cell strainer.
- Establishing a synchronous population
- Day -2: Tuesday (week 2), 1:00 p.m.
- Seeding the animals into plates
- Using a dissecting scope, check if the worms have hatched. Determine the concentration of worms in the S-complete buffer by counting the worm population in 10 µL drops using a dissecting scope; count 5-10 drops for each sample. If the concentration of worms in each 10 µL drop exceeds 30 worms per drop, dilute the total stock with S-complete to a lower worm density per drop to facilitate counting.
- Seed the liquid worm mixture into each well of a 96-well plate at a volume of 120 µL as follows (final concentrations): 60 worms/mL in S-complete, 50 µg/mL Carbenicillin, 0.1 µg/mL Amphotericin, and 6 mg/mL of OP50 prepared in section 1.
NOTE: The total volume to be prepared depends on the number of plates (~12 mL per 96-well plate). There should be 6-12 worms in each well. Alternatively, a large particle flow cytometry, such as the COPAS FP by Union Biometrica, can be used to precisely sort 10 worms into each well (See crowding effect section). - Also include a no-worms liquid mixture: 50 µg/mL of Carbenicillin, 0.1 µg/mL Amphotericin, and 6 mg/mL of OP50 prepared in section 1.
- Put 120 µL of the no-worms mixture into row H of the 96-well plate, which is used to determine the selflysis values, as mentioned earlier. Put 120 µL per well of the mixture with worms in rows A-G.
NOTE: Ensure the worms are kept in suspension during pipetting (see Figure 2A, B). We use clear 96-well plates with a flat bottom. Our plate layout divides the plate to produce 4 or 6 conditions in which each pair of columns will be a different drug treatment or worm strain, and the bottom row will serve as a "no worm" control (Figure 2A, B). - To avoid contamination and evaporation, seal the plate with a tape sealer. Incubate the plates for approximately 65 h at 20 °C until the animals become L4 worms.
- Seeding the animals into plates
- Day 0: Thursday (week 2) before noon.
- Sterilization of the worm population through the addition of Fluorodeoxyuridine (FUdR).
- Add 30 µL of a 0.6 mM FUdR stock solution to sterilize the animals in each well. Reseal the plate using tape sealers and shake the plates for 20 min on a microtiter plate shaker at 800 rpm. Return plates to the 20 °C incubator.
NOTE: In this step, the final OP50 concentration is reduced from 6 mg/mL to 5 mg/mL (1 × 109 bacteria/mL), and each well has a final volume of 150 µL. It is crucial to add FUdR to the L4 stage before the animals reach adulthood.
- Add 30 µL of a 0.6 mM FUdR stock solution to sterilize the animals in each well. Reseal the plate using tape sealers and shake the plates for 20 min on a microtiter plate shaker at 800 rpm. Return plates to the 20 °C incubator.
- Sterilization of the worm population through the addition of Fluorodeoxyuridine (FUdR).
- Day 1: Friday (week 2)
- Add drugs to the culture.
- By 9:00 a.m., confirm that most animals are gravid and each well contains several eggs. If necessary, add any drugs to the desired concentration (see the discussion).
- After adding the drug, seal the plates with tape sealer and shake the plates for 20 min at 800 rpm on a plate shaker.
NOTE: The shaking ensures that all bacterial clumps are dissolved without touching the sealer. Both bacterial clumps or liquid on the sealer will distort the OD600 reading. If liquid drops are on the sealer, briefly spin it for 10-20 s at 310 × g. Shake again on the shaker for 5 min and measure. - After 20 min of shaking on the microtiter plate shaker, remove the lid and the seal and measure the OD600 in a plate reader; this is the Day 1 OD600 measurement. Seal and return plates to a 20 °C incubator.
NOTE: Because bacteria settle quickly, the reading should occur within 10 min after shaking the plates. - Use an inverted microscope to check the population of worms for signs of contamination or toxicity if a drug has been added. Return the plates to the 20 °C incubator.
- Add drugs to the culture.
- Day 4: Monday (week 3)
- Take the Day 4 OD600 reading.
- Repeat the reading procedure from Day 1 (step 2.5.1.3) to get the Day 4 OD600 value. Before the OD600 measurement, shake the plates for 20 min at 800 rpm on the microtiter plate shaker.
- On an inverted microscope, with ideally a 2x objective, count the population of worms in each well and record it in a spreadsheet. Return the plates to the 20 °C incubator after counting.
NOTE: See a sample of our scoring sheet provided in Supplemental Material. - After the Day 4 reading, the food intake measurements are complete. If required, keep these plates for lifespan analysis.
NOTE: This protocol is compatible with Solis and Petrascheck31.
- Take the Day 4 OD600 reading.
3. Analysis of food intake data
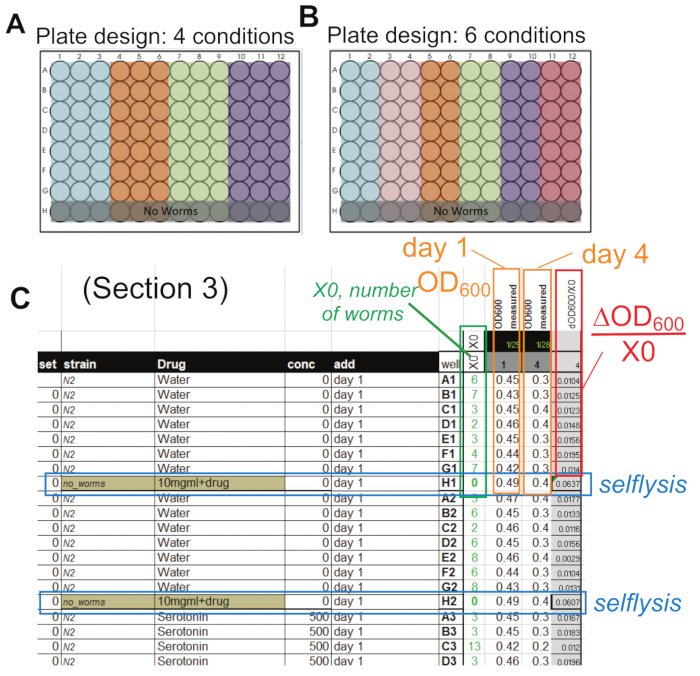
Figure 2: Design and analysis. (A, B) Possible plate designs for 4 and 6 conditions with the "no worm" control wells in row H. These wells are necessary to determine the selflysis rate. In each plate, always include an N2 untreated control population as a reference. (C) Sample data collected in the provided spreadsheet with the number of worms (green box), the two OD600 measures on days 1 and 4 (orange boxes), the selflysis values (blue box), and the evaluation using formula (2) ((OD600 –selflysis)/X0) in the red box. The specific example shown here uses the plate design shown in (B). Abbreviations: OD = optical density; X0 = number of worms per well. Please click here to view a larger version of this figure.
NOTE: This section describes how the food intake data from section 2 of this protocol are analyzed. The first values to be calculated are the selflysis control values. Bacteria lyse over time in liquid, even in the absence of worms. This selflysis value can also be affected by many chemicals and needs to be controlled as it is relatively large compared to a worm's food intake. As described in step 2.3.1.4 and shown in the plate layout in Figure 2A, B, row H contains the same solution, excluding the worms. In our plate setup shown in Figure 2B, there are 14 wells per condition with two respective selflysis control wells (row H, "no worms").
- Calculate the selflysis control values for the no-worms control wells in each condition using equation ( 1). For each replicate group of wells, ensure there are at least two no-worm control wells, as mentioned in step 2.3.1.4. Use the mean from all the no-worms control wells for the selflysis value for a replicate group.
Selfysis = mean(OD600 Day 1 – OD600 Day 4) (1)
NOTE: For the plate setup shown in Figure 2B, we would calculate six selflysis values, one for each of the six groups, by calculating the mean of the two selflysis control wells in row H. - Calculate the food intake per worm using equation
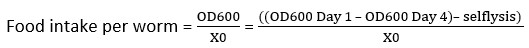 (2).
(2).
NOTE: Formula used in the spreadsheet (red box, Figure 2C) with X0 being the number of worms per well. - After the food intake per worm is determined for each well, calculate the averages and standard deviation for the entire condition.
NOTE: For the plate setup shown in Figure 2B, 14 replicate wells per condition are sufficient for statistical comparisons. - Normalize each condition to its respective N2 untreated control population. Indicate the change in feeding as a fold change or percentage of N2 feeding.
NOTE: Normalization is done because the absolute values of OD600/X0 from equation (2) can vary between experiments conducted on different days.
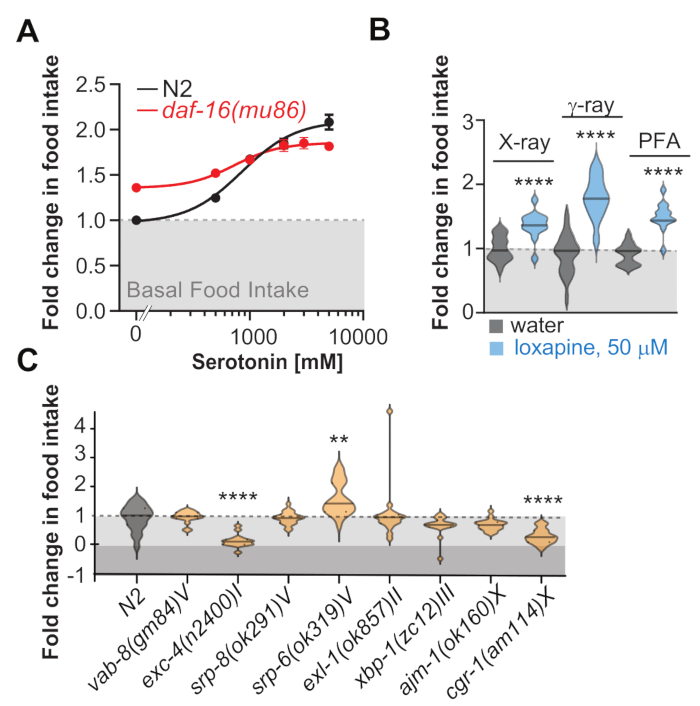
Figure 3: Representative data. (A) The graph shows dose-response curves of fold change in food intake as a function of serotonin concentration. All data show unstimulated basal food intake normalized to N2 basal feeding. Note that the daf-16(mu86) animals eat more than the wild type N2 when no serotonin was added but show a blunted range in their ability to control feeding. (B) Graph shows a violin plot of fold change in food intake of worms treated with 50 μM of the antipsychotic Loxapine but fed bacteria killed by either X-rays, γ-rays, or paraformaldehyde (C) Graph shows a violin plot of fold change in food intake of different mutants with their genotypes indicated in the x-axis. These data suggest that exc-4 and cgr-1 mutants eat less while srp-6 mutants eat more. Asterisks represent ** p < 0.001, ****p < 0.0001 as determined by ANOVA and Brown-Forsythe-Welch, post-hoc corrected to account for multiple hypotheses testing. Error Bars show ±S.E.M. Please click here to view a larger version of this figure.
Figure 1 highlights the overall workflow of this protocol. It illustrates the entire process, from making the bacteria to acquiring the OD600 reading on day 1 and day 4. In addition, the infographic references the specific steps of the methods being used. Figure 2A, B shows two possible experimental plate layouts, including "no-worms" control wells in row H, necessary for calculating the selflysis rates described in the discussion section. In our lab's experience, it is necessary for one of the conditions to be untreated N2 control to account for the variability of bacterial intake of worms observed between experiments (see discussion: Planning [N2 reference population]).
Figure 2C is an example of the spreadsheet used to analyze the data generated in this protocol. It highlights the worm count in green, selflysis de blue, day 1 OD600 and day 4 OD600 in orange, and the food intake per worm in red. In addition, each well is tracked for the strain, drug, concentration, date of experiment, and other treatments added. Further information on the specific equations used can be found in protocol section 3. Overall, these spreadsheets (template provided as Supplemental Materials) provide efficient data collection and analysis of this protocol.
Figure 3A shows representative results for a dose-response curve of how serotonin modulates feeding in N2 and daf-16 mutants using this protocol. In this experiment, the fold change from basal food intake (first dot in the curve) was calculated across five increasing doses of serotonin. As these results highlight, the N2 strain is able to overeat in a dose-dependent manner. However, for the daf-16(mu86) mutant, although basal feeding is higher than N2, the mutant cannot respond to serotonin in the same dose-dependent manner as the N2 strain. This protocol allows us to determine the feeding behavior between two strains where the concentration of serotonin was the variable factor to generate dose-response curves. Figure 3B shows the food intake of worms treated with the antipsychotic Loxapine but fed with bacteria killed by either X-rays, γ-rays, or paraformaldehyde. Note that these experiments were not conducted in parallel and that the differences are not representative of the different methods to kill the bacteria but are due to inter-experimental variability. Figure 3C shows the food intake of a series of genetic strains, with two strains showing a decrease and one strain showing an increase in feeding behavior relative to N2. This application of the protocol enables the testing of one drug at one concentration across different genetic backgrounds. It is suited to identify genetic backgrounds that show a different food intake response than wild-type N2 controls when given the same drug. It can be paired with our lifespan protocol to quickly screen food intake and lifespan in the same plate setup31.
Supplemental Material: Spreadsheet templates shown in Figure 2C. Please click here to download these materials.
